

Empfohlen
Weitere ähnliche Inhalte
Was ist angesagt?
Was ist angesagt? (20)
Ähnlich wie 130 anther dev, ovule dev.revised
Ähnlich wie 130 anther dev, ovule dev.revised (20)
Mehr von tralala 1412
Kürzlich hochgeladen
Kürzlich hochgeladen (20)
130 anther dev, ovule dev.revised
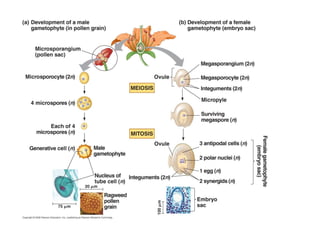
- 2. Microsporogenesis- formation of spores called microspores Microgametogenesis- development of microspore into the microgametophyte or the pollen grain containing sperm cells
- 3. Anthers surrounding a central ovary in Lily
- 4. microspores
- 12. Development of anther protoderm Hypoderm-found beneath protoderm and becomes Archesporial layer. Divides into:L 1.Pri parietal cells (outer)- differentiates into sporangial outer wall- --endothecium and tapetum 2. Primary sporogenous Sporangium initiation is restricted to four separated areas corresponding to corners of the developing anthers
- 13. Anther still meristematic lobe .Periclinal division occurs beneath the protoderm, layer called archesporial layer
- 14. At maturity, the 2 sporangia of each side become confluent due to breaking down of the partition between them Endothecium lying immediately beneath epidermis, hygroscopic- aids in dehiscence Microsporocytes-or microspore mother cells are derived from archesporial cells
- 16. Parts of ovule 1.nucellus- central body with vegetative cells enclosing the Sporogenous cells 2. 1 or 2 integuments (unitegmic or bitegmic) enclosing the Nucellus 3. funuculus-stalk connecting ovule with the placenta. 4. chalaza-where nucellus, integuments and the funiculus merge
- 17. X-section of an ovary Ovary contains a cavity lined with an epidermal layer. Ovules develop from the epidermal cells and are contained within the cavity of the ovary, attached to its inner surface by a short stalk- funiculus Ovary bears ovules on a ridge on the ovary wall called placenta
- 18. Developing ovules of Lilium. Ovule emerges from the placenta as conical protuberance with the first sporogenous cell,called archesporial cell. Integuments formed by periclinal div. of epidermis
- 19. Chalazal region
- 20. A, D. (bel mutant) exposed nucellus and a single integument F. Larger mutant ovule Outer integument has many cells.
- 21. Megaspore mother cell differentiates from surrounding nucellar tissue and undergoes meiosis. Inner integ. Outer integ Begin as ridges of tissue early in Ovule dev. C, E,F differential growth of ovule causes them to curve so micropyle is bent around funiculus and placenta (C).
- 22. Diakinesis- homologs are held together by chiasmata at their tips. Summary: ist meiotic prophase- replicated homologous chromosomes synapse, usually undergo crossing-over, then condense as tetrads. Held together at the centromeres, pairs of Sister chromatids in each tetrad are ready to be distributed to opposite .poles during the remainder of the first meiotic division
- 23. a,b) different stages of pachytene c.) metaphase with precocious migration to the pole d) Anaphase with laggard chromosome e) Telophase f) Metaphase II with precocious migration g) Late anaphase H) Telophase II i) tetrad
- 24. Tapetum- originates from Primary parietal cells. Dense cytoplasm, may become polyploid or multinucleate. completely invest the locule, have nutritional function by transferring food materials to the differentiating pollen grains
- 25. Meiosis I Pair and exchange segments Chromosomes line up by homologous pairs Each pair of homologous chromosomes separates Two haploid cells form, each chromosome still consists of two sister chromatids
- 26. Leptotene- chromatin condenses, preceded by DNA replication Zygonema-homologous chromosomes pair form bivalents Pachytene-physical exchange of chromosome parts occurs bet homologous chromosomes Diplotene- partial separation of each of sister chromatids from their homologous chromatids
- 27. Meiosis I Leptotene-chromatin condenses ,reveal individual chromosomes. Already replicated. Zygotene- separated homologous chromosomes pair (synapsis)with each other forming bivalents or chromosome pairs. Pachytene-Physical exchange of chromosome parts occur between homologous chromosomes. Arms of the two pairs of sister chromatids separate from each other and then each arm becomes closely associated with its homologous pair.tetrad- group of 4 chromatids Crossing-over or recombination occurs Diplotene- partial separation of each pair of sister chromatids from their homologous counterparts.Sister chromatids still held together. Diakinesis-chromatids condense and appear compact rods that are grouped as tetrads.
- 28. Chromosomes still composed of two chromatids Chromosomes at metaphase plate. Due to crossing – over in Meiosis I, each chromosome not genetically identical. Anaphase II Sister chromatids Separate, move to opposite poles as Individual chromosomes Telophase II and Cytokinesis. Nuclei form. Chromosomes begin decondensing Meiosis II
- 30. Meiosis Requires 2 nuclear divisions Produces 4 haploid daughter cells Replication occurs only once. Daughter cells receive on of each kind of parental chromosome but in different combinations. Daughter cells not genetically identical to parent cell Or to each other
- 31. Cells lining the anther lumen – a layer known as the endothecium – secretes materials that are essential for the proper maturation of the pollen grains. Cells of the endothecium are so densely cytoplasmic that no vacuoles at all are visible; it is even difficult to distinguish one cell from another in most of the endothecium. The dark red dots in the both the endothecium and the pollen grains are nucleoli, with the actual nuclei being just barely visible around a few of them
- 32. Anaphase of first mitosis Metaphase of 2nd mitosis
- 33. Telophase I cell plate will disperse without forming cell wall Early prophase i Mid prophase I Metaphase II
- 34. Typical anther has four elongated microsporangia. At maturity the two sporangia become confluent owing to the breaking down of the partition between them
- 35. tapetum
- 36. Pollen development and maturation The end of meiosis in the microsporocyte or microspore mother cell marks a turning point in microsporogenesis . Results in the production of 4 microspores, each with its own callose envelope. A candidate gene for separation of microspores from the tetrad in Arabidopsis anthers is designated as QUARTET (QRT). Outcome of microsporogenesis affected by this mutation is release of microspore in tetrads Failure of microspore separation in qrt mutants Traced to the fusion of exine layer of adjacent microspores Failure of protein degradation
- 37. Pectin is absent in primary wall of wild type microspores at the time of release from tetrad Pectin remains as integral part of the microspore wall of mutant QRT gene functions in degradation of pectin in order to separate the microspore from tetrad. Model of primary cell wall
- 38. In the wild types, after release from the tetrad, the microspore as the first cell of the gametophyte generation further differentiates to produce sperm cells. Internally, microspore increase in size and externally forms a double-layered wall, outer sculptured exine and inner smooth intine. First element of exine is called primexine- detected in the microspores while still in tetrad stage. Exine consists of sporopollenin, synthesized in the tapetum, substance resistant to chemicals and biodegradation. Exine formation is programmed by the diploid genome of microsporocyte. Intine is programmed by haploid genome of microspore, made up of cellulose microfibrils embedded in a matrix of pectin and hemicellulose
- 39. Pollen grain has two cells: from first mitotic division Vegetative cell- develops into pollen tube. Contains most cytoplasmic organelles Generative cell- small, produces the sperm -cytoplasm partitioned unequally during mitotic division of microspore -lacks mitochondria and chloroplast -at some point in pollen dev., divides by mitosis, each daughter cell differentiates into sperm cells, will lack also chloroplasts and mitochondria. This is the basis of for the maternal inheritance of chloroplast and mitochondrial genomes which occurs in ca 90% of all angiosperm species.
- 41. Development of embryo sac and female gamete (in an anatropous ovule) A hypodermal cell of the nucellus enlarges and becomes differentiated into a megaspore mother cell or megasporocyte. This diploid megaspore mother cell increases in size and undergoes meiosis to form a linear tetrad of 4 haploid megaspores, 3 of which degenerate and the 4th becomes the functional megaspore in monosporic types, all 4 become functional in tetrasporic types Female Gametophyte The nucleus of the megaspore undergoes three successive mitotic divisions forming eight nuclei. The megaspore enlarges into an oval shaped structure called the embryo sac. The eight nuclei of the embryo sac arrange themselves in 3 groups.
- 43. Micropyle Inner integument Outer integument placenta funuculus Outer and inner integument completely overgrow the nucellus Except for the micropyle. --Begins with elongation of the functional megaspore, usually at chalazal end. -- initially megaspore is non-vacuolate but later small vacuoles appear which may fuse to form large vacuole. Development of embryo sac
- 44. A. First megaspore mitosis yields binucleate embryo sac. Spindle of first nuclear div oriented along the long axis of the cell. Wall formation Does Not follow the nuclear division. Both nuclei divide 2x, forming 4 in B then 8 in C B. Large vacuole appears between the two daughter nuclei. As cell expands, nuclei are pushed toward opposite poles of the cell. Both nuclei from each pole divide twice
- 45. D. The 8 nuclei arrange themselves in two clusters of 4 nuclei one at each opposite ends. One nucleus from each end migrates towards the middle, called polar nuclei (named for where they came from, not where they end up). C.8-nucleate state . All 8 nuclei are present in a common cytoplasm, they move around probably from remnants of spindle fibers from earlier divisions.
- 46. Chalazal trio called antipodals ( Latin “against the foot”) at opposite end of the egg and antipodals Egg apparatus consists of larger egg flanked by two smaller cells called synergids (greek for “helpers” or cooperators The large binucleate
- 49. Megaspore mother cell devs. from surrounding nucellar tissue and undergoes meiotic division to form megaspore. Nucellus considered as a megasporangium funiculus nucellus chalaza- region where integuments fuse with funiculus
- 54. Mutants in ovule determination 1. bell (bel1)- ovule lacks inner integument 2. Aberrant testa shape (ats)- no clear distinction between inner and outer integument 3. Extreme types of integument mutations: aintegumenta (ant) huellenhos (hll) Do not develop integument and embryo is disrupted.
- 55. Megasporogenesis Differential growth causes ovules to curve so the micropyle is bent around to the funiculus and the placenta
- 56. Embryo sac cells 1.Egg- highly vacuolate, strongly polarized. In Arabidopsis, a large vacuole aligned toward micropylar end and an aggregation of cytoplasmic organelles and nucleus at chalazal end. Ultrastructural simplicity of cytoplasm characterize egg cells. amount of cytoplasm is limited cytoplasm spread as a thin layer surrounding vacuole cytoplasm contains very little ER, limited no. of plastids mitochondria, dictyosomes but high ribosomes which are randomly distributed rather than aggregated as polyribosomes cell wall does not extend over the entire cell but wall shows various attenuation toward chalazal pole
- 57. 2 Synergids- limited life span, wilt after fertilization. Probably involved in nutrition of egg. has extensive wall ingrowth at micropylar region called filiform apparatus metabolically active 3 Antipodals-transient existence , minimal cytoplasmic organelle show nuclear abnormalities like endoreplication 2 polar nuclei-metabolically active, extensive ER, numerous plastids, mitochondria, dictyosomes and polysomes, has large quantities of starch, proteins and lipids
- 58. Pollen grains
- 59. exine intine cytoplasm Pollen from different species, variation in exine morphology
- 60. Telophase of microspore mitosis in African lily. Most organelles are unequally segregated. Plastid is dividing adjacent to the chromatin of the future generative cell, but no plastids occur between cell plate and chromatin of the future generative cell. Cell plate Dividing plastid Generative cell Vege - tative cell
- 61. Post-meiosis: internal microspore/pollen events After a microspore enlarges in volume, unequal partitioning of cytoplasm takes place, it divides mitotically to form small lens to spheroidal shaped generative cell pressed against the vegetative cell membrane The generative cell moves away from the wall and into the interior of the vegetative cell after callose dissolves. Thus, one cell is completely surrounded by another cell. Generative cells typically become ovate to elongate while in the pollen grain. Lack plastids, before microspore mitosis, the plastids usually migrate to an area of the vegetative cell away from where the future generative cell will form.
- 62. A. Microspore B. Post-mitotic pollen grain with vegetative cell and newly-formed generative cell. C. Large central vacuole and generative cell appressed to wall
- 63. D. Pollen grain and generative cell have enlarged. E. Generative cell in mitosis F. Binucleate generative cell appressed to pollen wall
- 64. G. Two sperm cells still attached to each other but free from pollen wall; pollen engorging but central vacuole still present. H. Mature engorged pollen grain with separated lenticular sperm cells embedded in vegetative cell.
- 65. Plastids in generative cell or sperm cells are uncommon. No plastids in 18 grass species (includes common cereal grasses. None in any of the 7 crucifers (Brassicaceae) tested Among 39 legumes, 9 species had plastids. Pollen of most species shed from the anther with just generative and a vegetative cell. A sample of 2,000 dicots and monocots showed 30% were 3-celled
- 66. Landing on the stigma Pollen tubes
- 67. The exine protects the spore against dessication, mechanical pressure and ultraviolet radiation. Sometimes the exine layer is covered by sticky substances (pollenkitt, tryphine, elastoviscin and sporopollenin viscin threads), which are also produced by the tapetum. Pollenkitt- an adhesive material facilitates the attachment of pollen grains to insects, and in this way also zoophilic pollination. It also plays an important role in the adhesion of pollen grains to the female stigma and in the recognition between pollen and pistil. Also substances responsible for pollen allergy are often products originating from the tapetum. Pollen grains
- 68. Germination of pollen tube Pollen tubes extend up to sev. cm to reach embryo sac. Cell wall lacks cellulose but has another polysaccharide- callose- , a glucan Callose –synthesized by Golgi and transported to the extreme tip of pollen tube by Golgi-derived vesicles .Fusion of vesicles with plasma membrane expand the cell membrane of elongating tube Content of vesicles expand the wall of elongating tube
- 69. In angiosperms, to effect fertilization, the pollen grains germinate on the stigma by putting forth tubes (pollen tubes) which grow thru the style and find their way into the ovules where they discharge the sperms in the vicinity of the egg.
- 70. Page 114 Tube enters at the apex of the filiform apparatus and after growing thru it arrives in the cytoplasm of the synergid. The penetrated synergid starts degenerating before the arrival of the Pollen tube, but after pollination. The process of discharge takes place in seconds.
- 71. In cotton, the contents of the tube are discharged thru a subterminal pore which faces the chalaza.
- 72. Pollen tube discharge: includes 2 sperms, the veg, nucleus and a fair amount of cytoplasm. A portion of cytoplasm is retained in the pollen tube. No mixing between cytoplasm released by the pollen tube and that of the synergid. They remain as two separate entities.
- 74. Secretory vesicles originate from the Golgi network and are transported over the actin cytoskeleton into the growing tip where they fuse with the tip membrane expanding the wall and the plasma membrane. Besides pollen tube wall proteins, these transport vesicles contain mainly methyl-esterified pectins as part of the primary tube tip wall. Germinating pollen
Hinweis der Redaktion
- Figure 13.8 The meiotic division of an animal cell