2. Niveles estructurales en las proteínas
Estructura primaria: Secuencia de aminoácidos
Estructura secundaria: Plegamiento básico de la cadena
debido a enlaces de hidrógeno entre grupos -CO- y -NH-de
la unión peptídica: hélices, láminas y giros
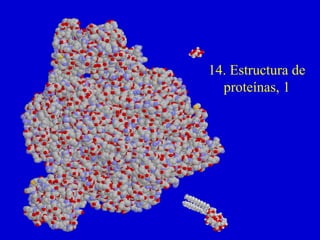
Estructura terciaria: Estructura tridimensional de la proteína
Estructura cuaternaria: Asociación de distintas subunidades,
siendo cada una un polipéptido.
4. Estructura primaria de la insulina
S S
GIVEQCCASVCSLYQLENYCN
S
S
S
S
FVNQHLCGSHLVEALYLVCGERGFFYTPKA
5. Oxidación de puentes disulfuro
H O
H
C
C
CH2
S
S
CH2
C
N
N C
H
H O
H O
H
C
C
CH2
N
SO3H
SO3H
CH2
C
N H
C
H O
HCO3H
6. H O
H
C
C
CH2
S
S
CH2
C
N
N C
H
H O
H O
H
C
C
CH2
N
DTT SH
ICH2 COOH
SH
CH2
C
N H
C
H O
H O
C
C
CH2
N
S
CH2
COO-H
COO-Reducción
CH2
S
CH2
C
N H
C
H O
y alquilación de puentes disulfuro
7. O2N
Determinación del N-término por la reacción de Sanger
F + H 2N C
NO2
N C
N
R1
O
H
R2
O
H
R3
O
O2N
HN C
NO2
N C
N
R1
O
H
R2
O
H
R3
O
O2N
HN
NO2
R1
COOH
R2 COOH
H2N COOH
R3
H2N
+ +
9. Rotura de un péptido por bromuro de cianógeno
O
N C
N C
N C
N
CH2
H
R
O
H
R
O
C C
N C
N
N C
H
O
R
H
O
R
H
H
R
O
H
R
O
CH2
S
CH3
N
N C
N C
H
O
R
H
O
R
H
O
O
C
N C
N
R
O
H
R
O
C
N C
H
R
O
H
R
O
BrCN
H2N
10. Degradación secuencial de Edman
N C
N C
O
H
R4
O
N C
C
H
O
R2
H
O
R1 R3
N C S + H2N
S
N C
N C
N C
O
H
R4
O
N C
C
H
O
R2
H
O
R1 R3
H
N
N C
N C
O
H
R4
O
Ácido diluído
H2N C
H
O
R2
R3
N
NH
S
O
R1
H
+
Feniltioisocianato
PTC-péptido
PTH-aminoácido
Péptido (n)
Péptido (n-1)
11. Hoy día, la mayor parte de estructuras primarias de proteínas
se determina a partir de la secuencia de nucleótidos en el genoma.
Técnicamente la secuenciación de ácidos nucleicos (en particular,
la de DNA) es mucho más sencilla y barata que la de proteínas,
estando al alcance de cualquier laboratorio.
12. 1 0 2 0 3 0 4 0 5 0 6 0
| | | | | |
P Y Q Y P A L T P E Q K K E L S D I A H R I V A P G K G I L A A D E S T G S I A K R L Q S I G T E N T E E N R R F Y R Q
7 0 8 0 9 0 1 0 0 1 1 0 1 2 0
| | | | | |
L L L T A D D R V N P C I G G V I L F H E T L Y Q K A D D G R P F P Q V I K S K G G V V G I K V D K G V V P L A G T N G
1 3 0 1 4 0 1 5 0 1 6 0 1 7 0 1 8 0
| | | | | |
E T T T Q G L D G L S E R C A Q Y K K D G A D F A K W R C V L K I G E H T P S A L A I M E N A N V L A R Y A S I C Q Q N
1 9 0 2 0 0 2 1 0 2 2 0 2 3 0 2 4 0
| | | | | |
G I V P I V E P E I L P D G D H D L K R C Q Y V T E K V L A A V Y K A L S D H H I Y L E G T L L K P N M V T P G H A C T
2 5 0 2 6 0 2 7 0 2 8 0 2 9 0 3 0 0
| | | | | |
Q K F S H E E I A M A T V T A L R R T V P P A V T G I T F L S G G Q S E E E A S I N L N A I N K C P L L K P W A L T F S
3 1 0 3 2 0 3 3 0 3 4 0 3 5 0 3 6 0
| | | | | |
Y G R A L Q A S A L K A W G G K K E N L K A A Q E E Y V K R A L A N S L A C Q G K Y T P S G Q A G A A A S E S L F V S N
H A Y Estructura primaria de la aldolasa A humana
SWISS-PROT http://www.expasy.ch
13. Cálculos a partir de estructura primaria
- Número, porcentaje y fracción molar de aminoácidos
- Fórmula molecular y peso molecular
- pI (punto isoeléctrico) teórico
- Absorbancia molar teórica
- Vida media teórica
- Índices de inestabilidad e hidrofobicidad
14. A m i n o a c i d c o m p o s i t i o n :
A l a ( A ) 4 2 1 1 . 6 %
A r g ( R ) 1 5 4 . 1 %
A s n ( N ) 1 4 3 . 9 %
A s p ( D ) 1 4 3 . 9 %
C y s ( C ) 8 2 . 2 %
G l n ( Q ) 1 7 4 . 7 %
G l u ( E ) 2 4 6 . 6 %
G l y ( G ) 3 0 8 . 3 %
H i s ( H ) 9 2 . 5 %
I l e ( I ) 2 0 5 . 5 %
L e u ( L ) 3 4 9 . 4 %
L y s ( K ) 2 6 7 . 2 %
M e t ( M ) 3 0 . 8 %
P h e ( F ) 8 2 . 2 %
P r o ( P ) 1 9 5 . 2 %
S e r ( S ) 2 0 5 . 5 %
T h r ( T ) 2 2 6 . 1 %
T r p ( W ) 3 0 . 8 %
T y r ( Y ) 1 3 3 . 6 %
V a l ( V ) 2 2 6 . 1 %
A s x ( B ) 0 0 . 0 %
G l x ( Z ) 0 0 . 0 %
X a a ( X ) 0 0 . 0 %
A t o m i c c o m p o s i t i o n :
C a r b o n C 1 7 4 1
H y d r o g e n H 2 7 8 0
N i t r o g e n N 4 8 6
O x y g e n O 5 2 6
S u l f u r S 1 1
F o r m u l a : C 1 7 4 1H 2 7 8 0 N 4 8 6O 5 2 6 S 1 1
T o t a l n u m b e r o f a t o m s : 5 5 4 4
M o l e c u l a r w e i g h t : 3 9 2 8 8 . 8
ProtParam, 1
http://www.expasy.ch
15. T o t a l n u m b e r o f n e g a t i v e l y c h a r g e d r e s i d u e s ( A s p + G l u ) : 3 8
T o t a l n u m b e r o f p o s i t i v e l y c h a r g e d r e s i d u e s ( A r g + L y s ) : 4 1
T h e o r e t i c a l p I : 8 . 3 9
E x t i n c t i o n c o e f f i c i e n t s :
C o n d i t i o n s : 6 . 0 M g u a n i d i u m h y d r o c h l o r i d e
0 . 0 2 M p h o s p h a t e b u f f e r
p H 6 . 5
E x t i n c t i o n c o e f f i c i e n t s a r e i n u n i t s o f M - 1 c m - 1 .
T h e f i r s t t a b l e l i s t s v a l u e s c o m p u t e d a s s u m i n g A L L C y s
r e s i d u e s a p p e a r a s h a l f c y s t i n e s , w h e r e a s t h e s e c o n d t a b l e
a s s u m e s t h a t N O N E d o .
2 7 6 2 7 8 2 7 9 2 8 0 2 8 2
n m n m n m n m n m
E x t . c o e f f i c i e n t 3 5 6 3 0 3 5 5 0 8 3 4 9 4 5 3 4 1 9 0 3 2 8 8 0
A b s 0 . 1 % ( = 1 g / l ) 0 . 9 0 7 0 . 9 0 4 0 . 8 8 9 0 . 8 7 0 0 . 8 3 7
2 7 6 2 7 8 2 7 9 2 8 0 2 8 2
n m n m n m n m n m
E x t . c o e f f i c i e n t 3 5 0 5 0 3 5 0 0 0 3 4 4 6 5 3 3 7 1 0 3 2 4 0 0
A b s 0 . 1 % ( = 1 g / l ) 0 . 8 9 2 0 . 8 9 1 0 . 8 7 7 0 . 8 5 8 0 . 8 2 5
ProtParam, 2
http://www.expasy.ch
16. E s t i m a t e d h a l f - l i f e :
T h e N - t e r m i n a l o f t h e s e q u e n c e c o n s i d e r e d i s P ( P r o ) .
T h e e s t i m a t e d h a l f - l i f e i s : > 2 0 h o u r s ( m a m m a l i a n r e t i c u l o c y t e s , i n v i t r o ) .
> 2 0 h o u r s ( y e a s t , i n v i v o ) .
? ( E s c h e r i c h i a c o l i , i n v i v o ) .
I n s t a b i l i t y i n d e x :
T h e i n s t a b i l i t y i n d e x ( I I ) i s c o m p u t e d t o b e 3 4 . 8 2
T h i s c l a s s i f i e s t h e p r o t e i n a s s t a b l e .
A l i p h a t i c i n d e x : 8 7 . 1 6
G r a n d a v e r a g e o f h y d r o p a t h i c i t y ( G R A V Y ) : - 0 . 2 6 8
ProtParam, 3
http://www.expasy.ch
17. Predicciones a partir de estructura primaria, 1
- Hidrofobicidad
- Estructura secundaria
- Retención cromatográfica en HPLC
- Residuos accesibles y ocultos
- Mutabilidad
18. A l a : 1 . 8 0 0
A r g : - 4 . 5 0 0
A s n : - 3 . 5 0 0
A s p : - 3 . 5 0 0
C y s : 2 . 5 0 0
G l n : - 3 . 5 0 0
G l u : - 3 . 5 0 0
G l y : - 0 . 4 0 0
H i s : - 3 . 2 0 0
I l e : 4 . 5 0 0
L e u : 3 . 8 0 0
L y s : - 3 . 9 0 0
M e t : 1 . 9 0 0
P h e : 2 . 8 0 0
P r o : - 1 . 6 0 0
S e r : - 0 . 8 0 0
T h r : - 0 . 7 0 0
T r p : - 0 . 9 0 0
T y r : - 1 . 3 0 0
V a l : 4 . 2 0 0
PYQYPALTPEQKKELSDIAH
Valor de hidrofobicidad para la
posición n (en este caso, 8):
n+4
bi = -10.5
S
i = n-4
Escala de hidrofobicidad
(según Kyte y Doolittle)
20. Predicciones a partir de estructura primaria, 2: Homologías
( C G 5 4 3 2 ) C G 5 4 3 2 P R O T E I N . [ D r o s o p h i l a m e l a n o g a s t e r ]
3 7 6 A A
S c o r e = 3 9 8 b i t s ( 1 0 1 1 ) , E x p e c t = e - 1 1 0
I d e n t i t i e s = 1 9 7 / 3 5 5 ( 5 5 % ) , P o s i t i v e s = 2 4 5 / 3 5 5 ( 6 8 % ) , G a p s = 2 / 3 5 5 ( 0 % )
Q u e r y : 2 Y Q Y P A L T P E Q K K E L S D I A H R I V A P G K G I L A A D E S T G S I A K R L Q S I G T E N T E E N R R F Y R Q L 6 1
+ Y P E + + E L I + + V A P G K G I L A A D E S + + K R Q I G E N T E E N R R Y R Q +
S b j c t : 5 F Y Y P - - N K E L Q E E L I C I S K A L V A P G K G I L A A D E S S A V M G K R F Q L I G V E N T E E N R R L Y R Q M 6 2
Q u e r y : 6 2 L L T A D D R V N P C I G G V I L F H E T L Y Q K A D D G R P F P Q X X X X X X X X X X X X X X X X X X P L A G T N G E 1 2 1
L T D + + I G V I + H E T L + Q + D D G P F + P L G + E
S b j c t : 6 3 L F T T D P K I A E N I S G V I F Y H E T L H Q R T D D G L P F V E A L R K K G I L T G I K V D K H F S P L F G S E D E 1 2 2
Q u e r y : 1 2 2 T T T Q G L D G L S E R C A Q Y K K D G A D F A K W R C V L K I G E H T P S A L A I M E N A N V L A R Y A S I C Q Q N G 1 8 1
T T Q G L D L + R C A Q Y K K + G F A K W R C + L K I + + T P S A I + E N A N V + A R Y A + I C Q
S b j c t : 1 2 3 F T T Q G L D D L A N R C A Q Y K K E G C S F A K W R C I L K I T K N T P S P Q A I L E N A N V M A R Y A A I C Q S Q R 1 8 2
Q u e r y : 1 8 2 I V P I V E P E I L P D G D H D L K R C Q Y V T E K V L A A V Y K A L S D H H I Y L E G T L L K P N M V T P G H A C T Q 2 4 1
+ V P I + P E + L G D H D L R C Q V E + L A V Y K A L S D H H + + L E G T L L + P + M V P G +
S b j c t : 1 8 3 L V P I I S P E V L A T G D H D L D R C Q K V N E I L L A G V Y K A L S D H H V F L E G T L L Q P S M V M P G L Q S N K 2 4 2
Q u e r y : 2 4 2 K F S H E E I A M A T V T A L R R T V P P A V T G I T F L S G G Q S E E E A S I N L N A I N K C P L L K P W A L T F S Y 3 0 1
+ I + A T V A + R R + V P P A V G + F G Q S E E E A + + + L N A I N P L K P W A + T F + +
S b j c t : 2 4 3 N H P P A D I G V A T V L A I R R S V P P A V M G V L F C G G A Q S E E E A T V H L N A I N N V P L C K P W A M T F A F 3 0 2
Q u e r y : 3 0 2 G R A L Q A S A L K A W G G K K E N L K A A Q E E Y V K R A L A N S L A C Q G K Y T P S G Q A G A A A S E S L 3 5 6
R A L Q S L + W G G K K E + A Q E + K R A N L A G K Y + A A + E L
S b j c t : 3 0 3 D R A L Q T S I L R T W G G K K E Q I S H A Q N E L I K R C R A N G L A S I G K Y V I G S V E S S A A T E R L 3 5 7
21. Dependiendo del grado de homología en su estructura
primaria, las proteínas se agrupan en:
- Superfamilias: homología en torno a 30 %
- Familias: homología superior a un 50 % y
la misma función, por lo general.
Además, hay pequeños tractos de secuencias comunes
a proteínas muy diversas, y que corresponden a ciertos
aspectos funcionales (como p.e. modificación
postraduccional): son los motivos secuenciales
22. Algunos motivos secuenciales en las proteínas
N-Glicosilación N-{P}-[ST]-{P}
Unión a glicosaminoglicano S-G-x-G
Fosforilación dependiente de cAMP [RK]-(2)-[ST]
Fosforilación, protein kinasa C [ST]-x(2)-[RK]
Fosforilación, tirosin kinasa [RK]-x(2)-[DE]-x(3)-Y
N-miristilación G- {EDRKHPFYW}-x(2)-[STAGCN]-{P}
Amidación C-terminal x-G- [RK]-[RK]
g-carboxilación de ácido glutámico x(12)-E-x(3)-E-x-C-x(6)-[DEN]-x-[LIVMFY]
Prenilación C-{DENQ}-[LIVM]
23. Predicciones a partir de estructura
primaria, 3: Filogenia y Taxonomía
- 8 1 1 0
_ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . G l y . _ _ _ . _ _ _ . _ _ _ . _ _ _ . G l y . _ _ _ . _ _ _ . _ _ _ . P h e .
2 0
_ _ _ . _ _ _ . _ _ _ . C y s . _ _ _ . _ _ _ . C y s . H i s . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . L y s . _ _ _ .
3 0 4 0
G l y . P r o. _ _ _ . L e u . _ _ _ . G l y . _ _ _ . _ _ _ . _ _ _ . A r g . _ _ _ . _ _ _ .G l y . _ _ _ . _ _ _ . _ _ _ . G l y . _ _ _ .
5 0 6 0
_ _ _ . T y r . _ _ _ . _ _ _ . A l a . A s n . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . T r p . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ .
7 0 8 0
_ _ _ . _ _ _ .T y r . L e u . _ _ _ . A s n . P r o . L y s . L y s . T y r . I l e . P r o . G l y . T h r . L y s . M e t . _ _ _ . P h e .
9 0 1 0 0
_ _ _ . G l y . _ _ _ . _ _ _ . L y s . _ _ _ . _ _ _ . _ _ _ . A r g . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ . _ _ _ .
1 0 4
_ _ _ . _ _ _ . _ _ _ . _ _ _ . Invariantes en citocromo c
24. Sustituciones conservadoras en citocromo c
P o s i c i ó n A m i n o á c i d o s
1 3 A r g , L y s
4 0 S e r , T h r
4 6 P h e , T y r
4 9 S e r , T h r
8 1 V a l , L e u , I l e
8 5 L e u , I l e
9 0 A s p , G l u
9 4 L e u , I l e
9 5 I l e , V a l
9 7 P h e , T y r
25. Sustituciones radicales en citocromo c
P o s i c i ó n A m i n o á c i d o s
3 3 H i s , S e r , T r p , T y r , A s n
4 4 A l a , P r o , A s p , G l n , V a l , G l u
5 4 A s n , A l a , S e r , G l n , A r g , L y s
6 0 G l u , G l y , A s p , A l a , G l n , L y s , A s n
6 5 T y r , P h e , S e r , M e t , A r g
8 3 P r o , A l a , V a l , G l y , T h r
8 8 P r o , A l a , A s p , G l u , L y s , T h r
8 9 G l n , L y s , A s n , G l u , T h r , G l y , A s p , A l a , S e r
9 2 A n a , A s n , A s p , G l y , G l u , V a l , T h r , L y s , G l n
26. Distancias filogenéticas en citocromo c
1 2 3 4 5 6 7 8 9
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
1 . H o m o s a p i e n s -
2 . M a c c a c a m u l a t a 1 -
3 . S u s s c r o f a 1 0 9 -
4 . G a l l u s d o m e s t i c u s 1 3 1 2 9 -
5 . R a n a p i p i e n s 1 8 1 7 1 1 1 1 -
6 . M u s c a d o m e s t i c a 2 7 2 6 2 2 2 3 2 2 -
7 . B o m b y x m o r i 3 1 3 0 2 7 2 8 2 9 1 4 -
8 . T r i t i c u m v u l g a r e 4 3 4 3 4 5 4 6 4 8 4 5 4 5 -
9 . N e u r o s p o r a c r a s s a 4 8 4 7 4 6 4 7 4 9 4 1 4 7 5 4 -
40. C
N C
N
N C
H O
O H
C
N C
N
H O
O H
C
N C
N
H O
O H
C
N C
N
C
N C
N
H O
O H
C
N C
N
H O
O H
C
N C
N
H O
O H
C
N C
N
C
N C
N
H O
O H
C
N C
N
H O
O H
C
N C
N
H O
O H
C
N C
N
C
C
C
N
N
N
Lámina b paralela
43. N C
C
H O H O H O
N C
N C
N C
N C
N C
N
H O H O H
O
H O H O H O
N
C N
C N
C N
C N
C N
C
O H O H O H
C
H O H O H O
N C
N C
N C
N C
N C
N
H O H O H
O
N
C
N
C
Lámina b antiparalela
52. Estructuras suprasecundarias
- Hélice-vuelta-hélice
- Siete hélices transmembrana y hélice anfipática
- Cremallera de leucina
- Unidad bab
- Meandro b
- Dedo de Zn
- Mano EF
61. Determinación experimental de la estructura secundaria
1. Métodos físicos:
Cristalografía Rayos X, Resonancia Magnética Nuclear
(RMN, NMR) en tanto en cuanto resuelven la estructura
terciaria
Otras técnicas: dicroísmo circular
2. Métodos predictivos a partir de la estructura primaria
62. Propensión de un aminoácido hacia una estructura dada
1. A partir de un conjunto de proteínas de estructura 3D conocida,
se forma la siguiente tabla:
Total a-hélice Estr. b Giro b
Glutamato 282 132 29 43
Aminoácidos 5507 1715 1555 1121
(Se ha puesto el Glutamato como ejemplo; esta tabla se prepara
para todos los aminoácidos)
63. 2. A partir de la tabla anterior, se calculan las frecuencias relativas
de aparición de dicho aminoácido en las tres estructuras:
a-hélice Estr. b Giro b
Glutamato 0.470 0.104 0.151
Aminoácidos 0.311 0.282 0.204
3. Se calcula entonces la propensión de cada aminoácido hacia una
estructura dada por el cociente de dividir la frecuencia relativa de
cada estructura por la frecuencia relativa media de todos los amino-ácidos.
En el caso del glutamato,
Pa = 0.470/0.311 = 1.511 Pb = 0.104/0.282 = 0.370
Pg = 0.151/0.202 = 0.748
64. A l a : 1 . 4 2 0
A r g : 0 . 9 8 0
A s n : 0 . 6 7 0
A s p : 1 . 0 1 0
C y s : 0 . 7 0 0
G l n : 1 . 1 1 0
G l u : 1 . 5 1 0
G l y : 0 . 5 7 0
H i s : 1 . 0 0 0
I l e : 1 . 0 8 0
L e u : 1 . 2 1 0
L y s : 1 . 1 6 0
M e t : 1 . 4 5 0
P h e : 1 . 1 3 0
P r o : 0 . 5 7 0
S e r : 0 . 7 7 0
T h r : 0 . 8 3 0
T r p : 1 . 0 8 0
T y r : 0 . 6 9 0
V a l : 1 . 0 6 0
PYQYPALTPEQKKELSDIAH
Valor de propensión hacia
a-hélice para la posición n
(en este caso, 8):
n+4
bi = 9.07
S
i = n-4
Escala de propensiones hacia a-hélice
(según Chou y Fasman)
65.
66. A l a : 0 . 8 3 0
A r g : 0 . 9 3 0
A s n : 0 . 8 9 0
A s p : 0 . 5 4 0
C y s : 1 . 1 9 0
G l n : 1 . 1 0 0
G l u : 0 . 3 7 0
G l y : 0 . 7 5 0
H i s : 0 . 8 7 0
I l e : 1 . 6 0 0
L e u : 1 . 3 0 0
L y s : 0 . 7 4 0
M e t : 1 . 0 5 0
P h e : 1 . 3 8 0
P r o : 0 . 5 5 0
S e r : 0 . 7 5 0
T h r : 1 . 1 9 0
T r p : 1 . 3 7 0
T y r : 1 . 4 7 0
V a l : 1 . 7 0 0
PYQYPALTPEQKKELSDIAH
Valor de propensión hacia
estructura b para la posición n
(en este caso, 8):
n+4
bi = 8.1
S
i = n-4
Escala de propensiones hacia estructura b
(según Chou y Fasman)
67.
68. A l a : 0 . 6 6 0
A r g : 0 . 9 5 0
A s n : 1 . 5 6 0
A s p : 1 . 4 6 0
C y s : 1 . 1 9 0
G l n : 0 . 9 8 0
G l u : 0 . 7 4 0
G l y : 1 . 5 6 0
H i s : 0 . 9 5 0
I l e : 0 . 4 7 0
L e u : 0 . 5 9 0
L y s : 1 . 0 1 0
M e t : 0 . 6 0 0
P h e : 0 . 6 0 0
P r o : 1 . 5 2 0
S e r : 1 . 4 3 0
T h r : 0 . 9 6 0
T r p : 0 . 9 6 0
T y r : 1 . 1 4 0
V a l : 0 . 5 0 0
PYQYPALTPEQKKELSDIAH
Valor de propensión hacia
giro b para la posición n
(en este caso, 8):
n+4
bi = 9.12
S
i = n-4
Escala de propensiones hacia giro b
(según Chou y Fasman)