1. ҚАЗАҚСТАН РЕСПУБЛИКАСЫ БІЛІМ ЖӘНЕ ҒЫЛЫМ МИНИСТРЛІГІ
ОҢТҮСТІК ҚАЗАҚСТАН ОБЛЫСЫНЫҢ БІЛІМ БАСҚАРМАСЫ
ШЫМКЕНТ АГРАРЛЫҚ КОЛЛЕДЖІ
«Генетика»
ОҚУ ҚҰРАЛЫ
Шымкент– 2016ж.
2. Дайындағандар: ауыл шаруашылық ғылымдарының кандидаты,
доцент Жолшыбек Тұрсынбек, оқытушы Шойбекова Құралай,
«Генетика» пәнінен оқу құралы. Шымкент
аграрлық колледжі.
1513000 – Втериария мамандығы студенттеріне «Генетика» пәніне
арналған оқу құралы. Осы оқу құралы жұмыстық оқу бағдарламасына
және күнтізбелік тақырыптық жоспарына сәйкес құрастырылған. Бұл оқу
құралында генетикалық талдау нәтижелері, будандастыру әдістері,
белгілер мен өзгергіштіктің тұқым қуалау заңдылықтарын зерттейтін
тақырыптар қамтылған.
Пікір жазғандар: ауыл шаруашылық ғылымдарының
кандидаттары, Б. Божбанов, М. Ермаханов
«Ветеринариялық жалпы кәсіптік пәндер» кафедрасының
отырысындақаралды. Хаттама № 3, 23. 12. 2016 ж
Шымкент аграрлық колледжінің оқу-әдістемелік кеңесінде талқыланып,
бекітілуге ұсынылды.Хаттама № __ «__» ________2016 ж
3. Мазмұны
Кіріспе.....................................................................................................................4
1-тақырып. Тұқым қуудың цитологиялықжәне биохимиялықнегіздері.........7
№ 1 Практикалық сабақ. Хромосомалардыңқұрылысыментанысу...............14
2-тақырып. Тұқым қуудың заңдылықтары. Жыныстықкөбейудің негіздері.
Менделизм.............................................................................................................17
№ 2 Практикалық сабақ. Моногибридті, дигибридтіжәне полигибридті
шағылыстырунәтижелерін талдау......................................................................23
№ 3 Практикалық сабақ. Дигибридтіжәне полигибридтішағылыстыру
нәтижелерін талдау. Белгілердің тәуелсіз ажырау заңы.................................
3-тақырып. Гендердің өзара әрекеттесуі............................................................31
№ 4 Практикалық сабақ. Гендердің өзараәрекеттесу кезіндегі тұқым
қуалауын талдау....................................................................................................36
4-тақырып. Тұқым қуудың хромосомалықтеориясы. Белгілердің тіркесіп
тұқым қууы............................................................................................................ 39
№ 5 Практикалық сабақ. Гендердің тәуелсіз алмасуы. Хромосомалардың
жұбынасипаттама беру........................................................................................44
5-тақырып. Жыныс генетикасы. Жыныспентіркескен белгілердің тұқым
қуалауы..................................................................................................................48
№ 6 Практикалық сабақ. Жыныспентіркескен белгілердің тұқым қуалауы.49
6 -тақырып. Микроорганизмдердіңгенетикасы. Ген инженериясы................52
№ 7 Практикалық сабақ. Ген тізбегін және рДНҚ тізбегін құрастыру...........59
7- тақырып: Иммуногенетика. Организмнiң иммундықжүйесi.....................61
№ 8 Практикалық сабақ. Қан тобынжәне ұрпақ генотипі арқылы әкесінің
генотипін анықтау................................................................................................66
8-тақырып: Өзгергіштіктің түрлері мен заңдылықтары. Тұқым қуалайтын
өзгергіштік............................................................................................................67
№ 9 Практикалық сабақ. Өзгергіштіктің түрлері. Модификациялық
өзгергіштік............................................................................................................71
№ 10 Практикалық сабақ. Тұқым қуатын мутациялардыңтүрлері. Тура және
кері мутациялар түрлері. Хромосомалықөзгергіштер....................................74
9-тақырып: Тұқым қуалайтын аурулар және олардыңалдын алу..................76
№ 11 Практикалық сабақ.Тұқым қуалайтын аурулар, олардыңалдын алу...78
Пайдаланылған әдебиеттер................................................................................79
4. 4
Кіріспе
Генетика организмдердiң тұқым қууы және өзгергiштiгi туралы ғылым.
Ол гректiң генезис – шыққан тегi деген сөзiнен шыққан.
Генетиканың мақсаты – тұқым қуу мен өзгергiштiктiң заңдарын ашу,
оларды басқару жолдарын анықтау, организмдердiң адамға қажеттi жаңа
түрлерiн шығару. Адамның, жанжануарлардың тұқым қуалайтын кеселдерiн
зерттеп, оларды генетикалық жолмен болдырмауды анықтау.
Тұқым қуу дегенiмiз – тiрi организмдердiң табиғи қасиеттерiн, белгiлерiн,
өздерiне ғана тән биосинтездi және зат алмасуды ұрпақтан ұрпаққа берiп
қайталап отыруы. Ал өзгергiштiк дегенiмiз ұрпақтардың ата-анасының
көшiрмесi болмай, олардан сәл де болса өзгеше болуы.
Генетиканың қысқаша даму тарихы және негізгі даму кезеңдері
1865 жылы чех жаратылыстану зерттеушісі Грегор Мендельдің "Өсімдік
будандарымен тәжірибелер" атты еңбегі жарық көрді. Өсімдіктерді
будандастыру бойынша қойылған ұқыпты тәжірибелер оған тұқым қуалаудың
негізгі заңдарын ашуға мүмкіндік берді. Осы зерттеулерінің нәтижесі 1865
жылы Брно (Словакия) қаласы табиғат зерттеушілер қоғамының отырысында
баяндалды. Бірақ, оның жұмысы өз замандастарының арасында толық қолдау
таппады, сонымен Мендельдің ілімі 35 жыл бойы ұмыт қалды.
1900 жылы Г.Д. Фриз (Голландия), К.Корренс (Германия), Э.Чермак
(Австрия) бір-бірімен байланыссыз, әртүрлі объектілермен тәжрибелер
жүргізіп, Мендель тапқан белгілердің тұқым қуалау заңдылықтарын қайтадан
ашты, сондықтан осы 1900 жыл генетиканың дүниеге келген, яғни туған жылы
деп есептелінеді.
Генетиканың даму тарихын шартты түрде негізгі бес кезеңге бөлуге
болады.
Бірінші кезең 1900-1912 жылдар аралығы. Бұл жылдар Мендель ашқан
тұқым қуалау заңдылықтарының беку жылдары болды. Әр түрлі елдерде,
түрлі объектілермен жасалған гибридологиялық тәжірибелерден орасан көп
деректер алынды, олар Мендель ашқан тұқым қуалау заңдарының
универсалды сипатта екендігін толық дәлелдеді. Аз жылдар ішінде генетика
өз алдына ғылым болып қалыптасты және кең көлемді ғылым ретінде
танылды. 1906 жылы ағылшын ғалымы У. Бэтсон жаңа ғылымның атын -
генетика деп атауды ұсынды (латынның geneo - шығу тегіне, тууға қатысты).
1909 жылы Дания ғалымы В. Иоғансен ген, генотип және фенотип деген
терминдерді ұсынып, оларды биологиялық әдебиеттерге енгізді. Голландия
ғалымы Г. Де Фриз 1901 жылы мутациялық өзгергіштік теориясынұсынды, ол
теория бойынша организмдердің тұқым қуалайтын қасиеттері мен белгілері
кенеттен өзгереді деген ұғым қалыптасты.
Екінші кезең Генетиканың дамуының келесі кезеңінде (шамамен 1912
5. 5
жылдан 1925 жылға дейін) тұқым қуалау факторлары
хромосомалармен байланысты екені дәлелденді. Тұқым қуалаудың
хромосомалық теориясын жасауда АҚШ ғалымы Т.Г. Морганның және
оның шәкірттерінің (А. Стертевант, Г. Меллер, К. Бриджес) жұмыстары
маңызды роль атқарды. Бұл авторлар, кейінірек генетикалық зерттеулердің
негізгі объектісі болған жеміс шыбыны дрозофиламен жасаған
тәжірибелерінде, гендердің хромосомаларда бірінен соң бірі тізбектеле
орналасатындығын, әр геннің ондабелгілі орны болатындығын және гендердің
бір-бірінен белгілі бір ара қашықтықта орналастындығын анықтады.
Тұқым қуалаудың хромосомалық теориясы биологияның ең ірі жетістігі
болды, бұл теория тек генетикалық мәліметтерге ғана емес, сол сияқты митоз
және мейоздағы хромосомалардың қозғалу сипатына, тұқым қуалаудағы
ядроның ролі туралы мәліметтерге де сүйенді. Генетиканың дамуының бұл
кезеңінде көптеген белгілі генетиктер құнды зерттеулер жүргізді.
Генетиканың көптеген проблемалары бойынша кең көлемде жүргізілген
зерттеулер нәтижесінде ол жылдары қысқа мерзім ішінде тұқым қуалауды
зерттеу саласында бұрынғы Кеңес үкіметі алдыңғы мемлекеттердің
қатарына шықты. Генетиканың дамуында Н.К. Кольцовтың, Ю.А.
Филипченконың, З.И. Вавиловтың, Г.Д. Карпеченконың,
А.С.Серебровскийдің т.б. ғалымдардың еңбектері маңызды роль атқарды.
Үшінші кезең Генетиканың дамуының үшінші кезеңінің (шамамен
1925-1940 жылдары) ең басты ерекшелігі - мутацияларды қолдан алу
мүмкіндігінің ашылуы. 1925 жылы орыс ғалымдары Г.А. Надсон мен Г.С.
Филиппов саңырауқұлақтармен, ал 1927 жылы АҚШ ғалымы Г. Меллер
дрозофиламен жасаған тәжірибелерінде ренттен сәулелерінің тұқым
қуалайтын өзгергіштіктердің пайда болуына себепкер екендігі туралы мәлім
етті. Кейінірек 30-40 жылдары химиялық қосылыстардың да мутациялар
тудыратыны анықталды (В.В. Сахаров, М.Е. Лобашев, И.А. Рапопорт). Бұл
кезеңде эволюцияның генетикалық негіздерін зерттеу бағытындағы
жұмыстар да дамыды (С.С. Четвериков, Р. Фишер, Дж. Холдейн, С. Райт).
Төртінші кезең шамамен 40-шы жылдардан 1955 жылға дейінгі уақытты
қамтиды. Бұл кезең биохимиялық және физиологиялық белгілер генетикасы
бойынша ғылыми жұмыстардың дамуымен сипатталады. Әртүрлі
организмдердің, соның ішінде дрозофила мен нейроспораның тұқымқуалайтын
белгі-қасиеттерінің қалыптасуы негізінде биохимиялық процестердің
жататындығын зерттеу геннің әрекетін түсіндіруге мүмкіндік туғызды. Дж.
Бидл және Э. Тэтум қандай болмасын ген организмде бір ғана ферменттің
түзілуін анықтайды деген қорытындыға келді. Содан келіп, "бір ген - бір
фермент", деген ұғым қалыптасты, кейінірек бұл ұғым "бір ген -бір белок",
"бір ген - бір полипептид" деген қағидаға ұласты. Микроорганизмдер
6. 6
гендерінің құрылымын молекулалық деңгейде талдауға мүмкіндік беретін
бірсыпыра жаңа генетикалық құбылыстар ашылды. Генетиканың даму
тарихындағы осы кезеңнің соңғы жылдарында бактерияларда
генетикалық трансформация мен трансдукция құбылыстарының
табиғаты анықталды. Вирустардың инфекциялық элементі олардың өз
нуклеин қышқылдары болып табылатындығы белгілі. Ф. Крик пен Дж.
Уотсонның 1953 жылы ДНҚ молекуласының құрылысын анықтауы аса зор
жетістік болды. Молекулалық биология мен молекулалық генетиканың
келешектегі көптеген бағыттарының дамуында ДНҚ-ның құрылысының
анықталуы орасан зор роль атқарды.
Бесінші кезең генетиканың қазіргі даму кезеңі (1955 жылдан осы
уақытқа дейін) тұқым қуалау құбылыстарын молекулалык деңгейде зерттеумен
сипатталады. Генетиканың дамуының осы кезеңінде ашылған жаңалықтарды
жай ғана санап айтудың өзі көп орын алар еді, дегенмен осы жетістіктердің
кейбіреулерін атап өтейіқ, олар: генетикалық кодтың анықталуы; геннің
химиялық синтезі; кері транскрипция құбылысының табылуы; гендердің экзон
- интрон құрылымының ашылуы; рекомбинанттық ДНҚ технологиясы;
генетикалық және клеткалық инжәнерия бойынша жұмыстардың
дамуы.
Қазақстанда молекулалық биология және гендік инжәнерия саласындағы
зерттеулер 60-шы жылдар аяғында басталды. Бұл зерттеулер ҚР ҰҒА-ның
құрамында молекулалық биология және биохимия институты ащылғаннан
кейін (1983) үдей түсті. Өсімдік клеткасындағы хромосомалар зерттеліп,
олардың бидай эмбриогенезі кезінде белок құрастыруға, бұл процесті реттеуге
қатысатыны анықталды. Өсімдік геномыныңмолекулалық құрылымы мен оның
экспрессиясы, клеткалық инженерия мен биотехнология мәселелері зерттелді.
Республика селекционерлері тұқым таңдау, жұптастыру, аса бағалы
генотиптерді іріктеп, селекцияда пайдалану тәсілдерін қолдана отырып, ақбас
сиырын, алатау, әулиеата сиырларын, қазақтың биязы жүнді қойын, оңтүстік
қазақтың мериносын, биязылау жүнді дегерес қойын, жүндес ешкіні, қостанай
жылқыларын, жетісу шошқасын шығарды.
Генетиканыңзерттеуәдістері
Тұқым қуалау мен өзгергіштікті зерттеуде бірқатар әдістер қолданылады.
Олардың ең негізгісі генетикалық талдау болып табылады.
Жынысты көбеюде организмдердің жеке қасиеттері мен белгілерінің
тұқым қуалауын және тұқым қуалау заңдылықтарын талдауға мүмкіндік
беретін будандастыру жүйесін сол сияқты гендердің өзгергіштігі мен олардың
комбинаторикасын зерттейтін әдісті гибридологиялық талдау деп атайды.
Бұл генетикалық талдаудың ішіндегі ең негізгісі. Бұл әдістің мәні бір немесе
бірнеше белгілері арқылы бір-бірінен ажыратылатын организмдерді
7. 7
будандастыру(гибридизация) болып табылады. Осындай будандастырулардан
алынған ұрпақтар гибридтер болып табылатындықтан, бұл әдіс
гибридологиялық әдіс деп те аталады. Сонымен, гибридологиялық талдау
генетиканың ең негізгі және арнайы әдісі болып табылады. Оған сонымен
қатар математикалық статистиканың элементтері де енеді.
Математикалық әдіс, әдетте, будандастыру бойынша жүргізілген
тәжірибелердің нәтижелерін өңдеу, белгілердің өзгергіштігін зерттеу және
зерттелген белгілер арасындағы байланыстарды табу үшін қолданылады.
Семьялардағы немесе туыс адамдар арасындағы тұқым қуалаушылыққа
талдау жасау арқылы олардың белгі қасиеттерінің ұрпақтарға берілуін
зерттеуге мүмкіндік беретін генетикалық әдістің бір түрін генеалогиялык деп
атайды. Бұл әдіс көптеген тұқым қуалайтын аурулардың себебін табуға
мүмкіндік берді.
Қорытындылап айтқанда генетика тұқым қуалаушылықты және тұқым
қуалайтын өзгергіштікті негізгі үш бағытта зерттейді: 1) организмнің көбею
процесіндегі гендердің жағдайы; 2) геннің материалдық құрылымы; және 3)
геннің онтогенез барысындағы өзгергіштігі мен қызметі (әсері).
Бақылау сұрақтары
1. Генетика пәні туралы түсінік беріңіздер.
2. Генетиканың негізгі мақсаты.
3. Генетиканың даму тарихына қысқаша анықтама беріңіздер.
4. Генетиканың негізгі зерттеу әдістерін атаңыздар.
1-тақырып. Тұқым қуудың цитологиялық және биохимиялық
негіздері.
Жасуша тіршіліктің негізі, яғни ол тірі материяның құрылымды
функционалдық байланысының универсалды бірлігі болып табылады.
Жануарлар, өсімдіктер, саңырауқұлақтар жасушаларының жалпы құрылысы
негізінен бірдей. Жасуша құрылысының жинақы схемасында құрамында
хромосомаларыбар ядросы және ішінде өзін-өзі ұдайы өндіретін органоидтары
- митохондриялары мен хлоропластары (өсімдіктерде) бар цитоплазмасы
болатыны көрсетілген. Олардан басқа, эукариоттар цитоплазмасында әр
уақытта тұрақты түрде болатын, бірақ өзін-өзі өндіруге қабілетсіз
органоидтарда болады. Оларға Гольджи аппараты, вакоульдар, лизосомалар
жатады.
Бактериялардың(прокариоттардың) клеткалары басқаша типте құрылған.
Оларда қалыптасқан клеткалық ядро болмайды. Ядроның орнына эукариоттар
клеткасының ядросына ұқсас - нуклеоидтары болады. Әр клетка плазмалық
мембранамен қоршалған. Плазмалық мембрана цитоплазманы қоршап тұрады
және клетка ішіндегі зат алмасуының реттелуінде аса маңызды роль атқаратын
клетканың функционалды бөлігі болып табылады. Клеткаға түсетін барлық
қоректік заттар және клеткадан шығарылатын бүкіл қалдықтар немесе секреция
өнімдері осы мембрана арқылы өтеді.
8. 8
Тыныштықта тұрған клеткалардан екі қабат ядролық
мембранамен қоршалған сфералық ядроны ажыратуға болады. Ядро көбінесе
клетканың ортасында орналасады. Бірақ кейбір клеткаларда ол еркін орын
ауыстыра алады және клетканың кез келген бөлігінде орналасуы мүмкін.
Ядро клеткада өте маңызды роль атқарады, ол клетка қызметінің
активтілігіне бағыт береді. Бөлінуі басталмаған клетканың ядросы
микроскоппен қарағанда біркелкі сияқты көрінеді. Ондай ядродан
қабықшаны, бір немесе бірнеше ірірек денешіктерді яғни - ядрошықтарды,
кейде тағы да бір ұсақ гранулаларды байқауға болады. Бірақ бөлінуге кіріспеген
кейбір ядролардан недәуір үлкен құрылымдар - хромосомаларды да табуға
болады. Хромосомалар -құрамында тұқым қуалау бірліктері немесе гендері бар
таяқша тәріздес денешіктер. Клеткалардың бөлінулері аралығында
хромосомалар ұзын, өте нәзік хроматин жіпшелеріне айналады. Оларды тек
электронды микроскоппен ғана көруге болады. Ядрошықтардың ішінде
рибосомалар түзіледі. Клетканың бөлінуге дайындалуы кезінде ядрошықтар
жойылады да кейіннен қайта пайда болады. Хромосомалардың құрамындағы
ДНҚ арқылы ядро клетканың дамуы мен оның тіршілік әрекеттерін басқаруда
маңызды қызмет атқарады.
Ядрода тек белгілі бір клетканың белгілері мен қасиеттерінің немесе ол
кетканың ішінде жүретін тіршілік процестерінің (мысалы, белоктың
синтезделуі) ғана емес, сол сияқты организмнің барлық белгілері туралы да
тұқым қуалау ақпараты сақталады. Ақпарат - хромосомалардың құрамына
кіретін оның ең маңызды бөлігі - ДНҚ молекулаларында жазылған. Клетка
тыныштық күйде түрғанда (яғни, клеткалардың екі бөлінулері аралығындағы
кезеңде) хроматиндер ядродашашырап жатады. Клетканың бөлінуге дайындалу
процесінде хроматин жинақталып тығыздалады, сөйтіп, хромосомалар түзіледі,
содан соң пайда болған хромосомалар бөлінген екі жаңа клеткаға бірдей
тарайды.
Жасушаның бөлінуі екі кезеңнен тұрады, оның бөлінуге дайындық кезеңі
– интерфаза, ал бөліну кезеңі – митоз деп аталады. Бөлінуге кіріскен кез келген
клетка бірқатар өзгерістерге ұшырайды, сол өзгерістерден клеткалық циклді
құрайды. Клеткалық цикл төрт кезеңнен тұрады: синтезге дейінгі (G1), ДНҚ
синтезі (S), синтезден кейінгі (G2) және митоз (М). Көпшілік организмдер үшін
клеткалық циклдің ұзақтығы 10-нан 50 сағатқа дейін болады. Митоздың дәл өзі
клеткалық циклдің 1/7 - 1/10 бөлігінен аспайтын уақытта өтеді. Мысалы:
сүтқоректілерде М- 1,0-1,5 сағатқа, G1 - 8-12 сағатқа, G2 - 2,0-5 сағатқа, S - 6-10
сағатқа созылады.
Көп клеткалы организмдердің бәрінде де митоздық бөліну жүреді. Митоз
ядроның өзінің бөлінуі - кариокинезден және цитоплазманың бөлінуі -
цитокинезден тұрады. Клетка бөлінулерінің арасындағы кезең интерфаза деп
аталады. Интерфаза мен митоз клеткалық циклді құрайды. Митоз барысында
клетка бірқатар өзгерістерге ұшырайды, ол өзгерістер бірнеше фазаларға
бөлінеді: профаза, метафаза, анафаза және телофаза.
Клетка бөлінбей тұрған кезде, яғни интерфазада, метаболитгік процестер
интенсивті түрде жүріп жатады. Соның негізінде клетканың өсуі мен оның
келесі бөлінуіне қажетті әр түрлі заттар синтезделеді.
9. 9
Интерфазаның G1 - кезеңі
Интерфазаның G1 - кезеңінде клетка тіршілігі үшін аса қажетті заттар -
нуклеотидтер, аминқышқылдары, ферменттер т.б. синтезделеді. Бұл ең ұзақ
кезең, сонан соң ДНҚ синтезі жүретін - S - кезеңі келеді. Клеткадағы ДНҚ
мөлшepi екі есе артады. Осы уақыт ішінде басқа да заттар - РНҚ және белоктар
синтезделеді. Синтезден кейінгі G; - кезеңінде РНҚ және басқа да заттар
синтезделе береді. Бұл кезеңде клетка келесі митозға қажетті энергия
жинақтайды. Осы кезеңде ДНҚ-ның жаңадан түзілген тізбектерінің негізінде,
клеткада әр хромосоманың екі еселенуі басталады. Интерфаза негізінде
хромосомалар активті түрде қызмет жасайды, және микроскоппен қарағанда
олар көрінбейді. Митоздың профазасында хромосомалар ширатылады,
қысқарады және жуандайды. Сондықтан осы кезеңде олар бір центромера
арқылы біріккен екі хроматидтен тұратын құрылым ретінде көрінеді.
Хромаатидтер интерфазадағы хромосомалардың екі еселену процесінде пайда
болады. Профазаныңсоңына қарай ядрошықтар жойылады, ядролық мембрана
ериді. Сөйтіп хромосомалар цитоплазмада жатқандай болып шығады. Клеткада
бөлінудің арнайы механизмі болады. Профазаныңбасында хромосомалар бүкіл
ядро бойыншабіркелкі таралады, ал профазаның соңында ядролық мембранаға
жақындайды. Жіпшелердің күшті ширатылуына байланысты хромосомалардың
ұзындығы недәуір қысқарады.
Профаза. Митоздың профазасында хромосомалар ширатылады,
қысқарады және жуандайды. Сондықтан осы кезеңде олар бір центромера
арқылы біріккен екі хроматидтен тұратын құрылым ретінде көрінеді.
Хроматидтер интерфазадағы хромосомалардың екі еселену процесінде пайда
болады. Профазаныңсоңына қарай ядрошықтар жойылады, ядролық мембрана
ериді. Профазаның басында хромосомалар бүкіл ядро бойынша біркелкі
таралады, ал профазаның соңында ядролық мембранаға жақындайды.
1-сурет. Митоз схемасы. Клетка бөлінуі кезіндегі әртүрліфазадағы ядро:
10. 10
а-интерфаза; б-алғашқы профаза; в-ортаңғы профаза;г-соңғы профаза;
д- метафаза; е-анафаза; ж-телофаза; з-соңғы телофаза; и-екіге
бөлінген клетка.
Метафаза. Ядролық мембрана еріген соң хромосомалар экватор бағытына
қарай жылжиды. Митоздың бұл стадиясы прометафаза деп аталады.
Хромосомалардың жылжуы ахроматин жіпшелері (ұршық тәрізді) арқылы
жүзеге асады. Барлық хромосомалардыңцентромералары экватор жазықтығына
жеткен кезде центромераучаскелеріне бекіген клетка метафаза стадиясына
көшеді деп айтуға болады.
Егер осы стадиядағы клетканы микроскоппен қараса, онда барлық
хромосомалардың бір жазықтықта жатқанын және экваторлық немесе
метафазалық пластинка деп аталатын құрылым түзетінін жақсы көруге
болады. Осы стадияда клеткадағы хромосомалардың санын есептеу, олардың
құрылымын зерттеу және мөлшерін анықтау жұмыстары оңай жүргізіледі.
Метафазада байқалатын хромосомалардың саны мен пішіні түрдің кариотипін
сипаттайды.
Митоздың келесі фазасы - анафаза бұл центромера учаскесінің
бөлінуімен сипатталады, осы бөлінудің нәтижесінде хроматидтер
хромосомаларға айналады. Екі полюстерден келетін ұршық тәріздес жіпшелер
хромосомаларды қарама-қарсы полюстерге тарта бастайды. Клетканың
профазасында қанша хромосома болса, анафазаның соңында әр полюстерде
соншама хромосомадан болады.
Телофаза - митоздың ақырғы стадиясы бұл хромосомалардың
полюстерге толық ажырауымен бітеді. Профазада болатын барлық процестер
телофазада да қайталанады, бірақ олар керісінше жүреді, яғни ядролық
мембрана түзіледі, ядрошықтар пайда болады, хромосомалар тарқатылады
(деспирализация) және олар жіңішкеріп ұзара бастайды. Микроскоппен
қарағанда қайтадан тек тор құрылым көрінеді. Осымен ядроның бөлінуі –
кариокинез аяқталады.
Хромосомалардың құрылысы және функциясы
Әрбір ядрода хромосомалардың тұрақты саны және олардың
әрқайсысыныңөзіне тән генетикалық мәні болады. Әрбір юмосома хромонема
деп аталатын ДНҚ жіпшесінен тұрады, ол жіпшенің бойында тізілген моншақ
сияқты құрылымдар - хромомерлерорналасқан. Хромосоманың міндетті түрде
болатын құрылым ерекшелігіне бірінші жинақталу жатады, ол хромосоманы екі
бөлікке бөледі. Бірінші жинақталудың ішінде ерекше түзіліс - центромера
болады, ол хромосомалардың митоздық бөлінуінде маңызды роль атқарады.
Центромера профазадағы хромосомалардың пішінін анықтайды.
Хромосомалардың әр жұбының біреуі аналық, екіншісі аталық
организмнен келеді. Әртүрлі организмдердің сомалық тканьдері
клеткаларындағы хромосомаларды зерттеу тканьдердің тек өзіне ғана тән
хромосома болатынын көрсетгі. Әр түрге тән хромосомалар жиынтығының
ерекшеліктері – хромосомалардың саны және формасы (пішіні) оның
кариотипі деп аталады.
11. 11
Кариотиптегі хромосомалар саны жануарлар мен өсімдіктердің құрылым
деңгейімен байланысты емес. Жоғары сатыдағы организмдерге қарағанда
қарапайым организмдердің хромосомалар саны көп болуы мүмкін.
Хромосомаларды жұбымен топтастыра отырып, профазалық немесе
метафазалық хромосомалардың микрофотографиялары бойынша идиограмма
(хромосомаларды ұзындығы бойынша орналастыру) құруға болады. 79-шы
жылдардың басында дифференциалды түс әдісі хромосомаларды бір-бірінен
ажыратуға мүмкіндік берді. дрозофиланың), ішек клеткаларының, мальпиги
түтікшелерінің клеткалары -ядроларынан табыдды. Алып хромосомалар, сол
сияқты кейбір өсімдіктер синергидтерінің ядроларынан, қарапайым
жануарлардан да табылды. Интерфазалық сомалық және жыныс
клеткаларындағы хромосомаларға қарағанда, алып хромосомалар олардан 100-
200 есе ұзын және 1000 есе жуан. Алып хромосомаларды қарағанда, олардан
көлденең жолақтар көрінеді. Ондай жолақтар ашығырақ және күңгірттеу -
дискілердің кезектесуінен түзіледі. Ол жолақтар - дискілер бір-біріне тығыз
жанасқан, күшті ширатылған хромонемалар учаскелері немесе хромомерлер
болып табылады. хромомерлер политенді хромосомалар деп аталады.
Дискілердің көлемі мен морфологиясы өте өзгергіш келеді, бірақ әр хромосома
үшін олар тұрақты және оны ұқсастыру (идентификациялау) үшін маркер
қызметін атқара алады. Алып хромосомаларға тән аса үлкен көлемділік пен
пішіндер, олардың барынша көп тарқатылуы және хромосомалардың ажырап
кетпей ұдайы өзін-өзі өндіруі нәтижесінде пайда болады. Ядро мен клетканың
бөлінуінсіз-ақ, өзін-өзі өндіру есебінен хромосомалар санының көбеюін
эндомитоз деп атайды. Осының арқасында өндірілген хроматидтер ажырап
кетпейді және бір-біріне тығыз жанасып қала береді.
Мейоз. Клеткалар митоздық жолмен бөлінгенде хромосомалардың
тұрақты саны сақталады. Онда да, бастапқы және жаңа екі клеткалардағы
хромосомалар жиынтықтары өзара ұқсас болады. Егер жыныс клеткаларының
түзілуі де осындайжолмен жүрсе онда ұрықтанғаннан кейін хромосомалардың
саны әруақытта еселеніп артып отырған болар еді. Мейоз нәтижесінде
гаметалар - жұмыртқа клеткалары мен сперматазоидтар яғни жыныс
клеткалары пайда болады. Гаметалар түзілуі кезінде редукциялану, яғни
хромосомалар санының екі есе азаю процесі жүреді. Редукциялық бөліну
жануарларда гаметалар түзілуі кезінде (генетикалық редукция), өсімдіктерде
споралар (споралық редукция) түзілуі кезінде байқалады. Мейоз нәтижесінде
пайда болған гаметаларда хромосомалардың бір ғана, яғни гаплоидты
жиынтығы (n) болады.
Мейоз барлық организмдерде дерлік ұқсас жолмен жүреді. Мейоздың екі
бөлінуі шартты түрде бірінші мейоз (мейоз I), редукциялық және екінші мейоз
(мейоз II), эквациялық деп аталады. Митоз сияқты мейоздық бөліну де
профаза, метафаза, анафаза және телофаза стадияларынан тұрады. Мейоздың
алдында интерфаза процесі, ал ондахромосомалар редупликациясы - ДНҚ-ның
синтезі жүре
Редукциялы бөліну. Редукциялы бөліну митоз сияқты төрт фазадан
тұрады; профаза, метафаза, анафаза және телофаза. Редукциялы бөлінудің
профазасы өте күрделі. Ол бірін-бірі толықтыра жүретін бірнеше сатыдан
12. 12
тұрады. Атап айтқанда, жіңішке жіпшелер сатысы, қосарланған жіпшелер
сатысы, және жіпшелердің бір-бірінен алыстау сатысы және т.б. деп аталады.
Жіңішке жіпшелер сатысы (лептотена). Бұл сатыда хромосомалардың
жіпшелері екі еселеніп шумақ түзіледі. Электрондық микроскоппен қарағанда
әрбір хромосоманың екі жіпшеден тұратындығы байқалады. Бұл – хромосома-
лардың екі еселенуінің профазада жүретіндігінің дәлелі.
Қосарланған жіпшелер сатысы (зиготена). Бұл сатыда 2-суретте
көрсетілгендей, гомологті хромосомалар бір-біріне тартылып жақындасады.
2-сурет. Ұрық клеткаларының жетілу барысындағы мейоздық бөлінуі:
а-лептотена стадиясы; б-зиготена стадиясы; в-пахитена стадиясы; г-
диптена стадиясы; д-диакинез стадиясы; е-метафаза; ж-анафаза; з-те-
лофаза; и-мейоздың екінші бөлінуіндегі профаза; к- екінші бөлінудегі
метафаза; л- екінші бөлінудегі анафаза; м- екінші бөлінудегі телофаза;
Хромосомалардыңөзаражақындасуынконъюгация (жабысу) деп атайды.
Хромосомалардың конъюгациялануы оның ұш жағынан басталып, ортасына
қарай ауысса, ал екінші бір хромосомада конъюгация процесі олардың
ортасынан ұштарына қарай ауысатандығы дәлелденді. Осыдан кейін
хромосомалар ширатылып, өзіне тән пішіні мен мөлшеріне келеді. Демек, ол
ұзынынан қысқарып, көлденеңінен жуандайды
Жіпшелердің бір-бірінен алыстау сатысы (диплотена). Бұл сатыда
конъюгация барысына қарама-қарсы процесс жүреді, яғни жақындасқан
хромосомалар бір-бірінен ажырайды (2-сурет). Ұқсас (гомологті) хромосомалар
бір-бірінен алшақтағанда, Х тәрізді фигуралар пайда болады, олар хиазмалар
13. 13
деп аталады. Хиазмалар кезінде ұқсас хромосомалар өздерінің үлескілерімен
алмасады, мұны айқасу немесе кроссинговер деп атайды. Ал аналық-аталық
хромосомалар өзінді бөліктерімен алмасқанда, тұқым қуалайтын өзгергіштік
пайда болады. Осы кезде хромосомалардың санын анықтап, пішіні туралы
мағлұматтар алуға болады. Соңынан хромосомалар кариоплазмаға ретсіз
таралады, бұл – метафазаның басталғанының белгісі.
І-ші метафазада биваленттер метафазалық пластинканы құра отырып,
клетканың экватор жазықтығына орналаса бастайды. Хромосомалар толық
жуандайды және қысқарады. Организмнің сомалық клеткасындағы
хромосомалар санынан биваленттер саны екі есе кем, яғни гаплоидты санға тең
болады.
І-ші анафазада бір центромераға бекіген екі хроматидтерден тұратын
хромосомалар қарама-қарсы полюстерге таралады. Митоздың анафазасынан
мейоздың І-ші анафазасының басты айырмашылығы осы болып табылады. Әр
биваленттің аталық және аналық центромералары қарама-қарсы полюстерге
тарайды және олар бір-біріне тәуелсіз қозғалады. Одан әрі қарай
центромералардың редукциясы жүреді.
І-ші телофаза ядролықмембрананыңтүзілуімен және ядро құрылымының
қалпына келуімен сипатталады. Содан кейін қысқа стадия интерфаза немесе
интеркинез басталады. Кәдімгі интерфазадан интеркинездің айырмашылығы
сол, интеркинезде хромосомалар екі еселенбейді және ДНҚ синтезі жүрмейді.
3-сурет. Жыныс жасушаларының дамуы. Сперматогенез (А) және
овогенез (Ә); 2n - хромосомалардың диплойдты жиынтығы, n -
хромосомалардың гаплойдты жиынтығы.
Интеркинездің ізінше мейоздың екінші бөлінуі - эквациялық бөліну
басталады. Ол бөліну митоз типімен жүреді.
ІІ-ші профазада хромосомалардың ширатылуы есебімен олар жақсы
көріне бастайды. Ядролық қабықша, ядрошық жойылады, ұршық тәрізді
жіпшелер түзіледі.
ІІ-ші метафазада барлық хромосомалардың центромералары экватор
жазықтығына орналасады. Полюстен қараған кезде, клеткалардың
14. 14
хромосомалар саны гаплоидты, ал бірақ әр хромосома екі хроматидтерден
тұратындығы анық көрінеді.
ІІ-ші анафазада екі еселенген центромералар бір-бірінен ажырайды
нәтижесінде жаңа түзілген хроматидтер әр полюстерге тарайды.
ІІ-ші телофазада гаплоидты төрт ядролар түзіледі. Содан соң цитокинез
жүреді де, нәтижесінде төрт клетка пайда болады.
Сонымен, екі бөлінуден (редукциялық және эквациялық) тұратын мейоз,
бастапқы клеткаға қарағанда хромосомалар саны екі есе кем болатын төрт
клетканың пайда болуын қамтамасыз етеді.
Бақылау сұрақтар:
1. Митоз қандай фазалардан тұрады?
2. Митоз фазаларында қандай өзгерістер болады?
3. Меоз кезінде хромосомалардың санында қандай өзгерістер болады?
4. Редукциялы бөлінуге түсінік бер және оның фазаларын ата?
5. Эквациялық бөліну дегеніміз не?
№ 1 Практикалық жұмыс
Хромосомалардың құрылысымен танысу
Сабақтың мақсаты: Хромосомдар құрылысымен танысу. Кариотипті,
диплоидты, гаплоидты хромосомолар жиынтығын анықтау. Митоз және мейоз
кезіндегі хромосомалардың ерекшеліктерімен танысу. Жұп хромосомалардың
редукциялық бөлінуін зерттеу.
Құрал-жабдықтар: Әдістемелік нұсқау карталары, суреттер мен
кестелер, оқулық «Генетика».
Хромосома клетка ядросының генетикалық информация сақаталатын
құрылымы. Ол гректiң хромо- бояу, сомо дене деген сөздерiнен шыққан.
Интерфаза кезiнде клеткадағы хромосома-лар көрiнбейдi. Оларды тек
клеткалар бөлiну процессiкезiнде ғана көруге болады. Сондықтандаоны кейде
митотикалық хромосомалар деп атайды. Хромосомалардың көлемi әртүрлi:
0,2 мкм-нен 50 мкм-ген дейiн, диаметрi 0,2 мкм-нен 3 мкм-ге дейiн жетедi.
Әрбір ядрода хромосомалардың тұрақты саны және олардың әрқайсысының
өзіне тән генетикалық мәні болады. Хромосоманың міндетті түрде болатын
құрылым ерекшелігіне бірінші жинақталу жатады, ол хромосоманы екі бөлікке
бөледі. Бірінші жинақталудың ішінде ерекше түзіліс - центромера болады, ол
хромосомалардың митоздық бөлінуінде маңызды роль атқарады. Центромера
прафазадағы хромосомалардың пішінін анықтайды. Егер центромера
хромосоманың ортасында орналасса және хромосоманы тең екі иыққа бөлсе,
онда хромосоманың құрылымы X- тәріздес болады, оны метаорталықты
хромосома деп атайды. Центромера хромосоманы тең екі иыққа бөлмесе, ондай
хромосомаларды акроцентрлі хромосомалар деп атайды. Егер центромера
хромосоманың ұшына өте жақын орналасса және хромосома иықтарының
15. 15
біреуі ғана анық көрінсе, оны телоорталықты хромосома деп атайды. Кейбір
хромосомалардың ішінде екінші бунақталу болады. Кейде екінші бунақталудың
көрінуі қаншалықты анық, хромосома иықтарының біреуінің шеткі бөлігі ол
хромосома мен тек жіңішке жіп арқылы ғана жалғасқан сияқты болып көрінеді.
Ондайхромосома«серікті» (қосшы) хромосомадепаталады.
4 - сурет. Хромосомалардыңметафаза кезіндегі пішіні.
1- метаорталықты хромосома;2 -субметаорталықты; 3- акроорталықты; 4– тело-
орталықты; 5 - метаорталықты серігімен (SAT - хромосома); а-серігі; б - иығы;с-
центромера.
Хромосомалардың толық жиынтығын диплойдты, оның жартысын
гаплойдты деп атайды. Диплойдты хромосомалар жиынтығы дене (сомо)
жасушаларында болады. Гаплойдты хромосомалар жиынтығы жыныстық
жасушаларда болады. Дене (сомо) жасушалары митоздық жолмен бөлінеді, ал
жыныстық жасушалар мейоздық жолымен бөлінеді.
Жыныс хромосомаларынан басқа аутосомалық хромосомаларда жұп
хромосомалар морфологиялық құрылысы жағынан бiр-бiрiне ұқсас келедi.
Ондай хромосомаларды гомолог хромосомалар деп атайды. Жыныс
хромосомалары жұбынекi түрлi хромосомалар түзедi. Жыныс хромосомалары
екеуi бiрдей болса, оны гомогаметалы организмдер деп атайды.
16. 16
1-кесте.
Жануарлар мен өсімдіктердің соматикалық клеткаларындағы
хромосомалардың саны
Адам мен үй
жануарлары
Хромосома-
лардың саны
Өсімдіктер Хромосома-
лардың саны
Адам 46 Қатты бидай 42
Жылқы 66 Сұлы 42
Мүйізді ірі қара
мал
60 Картоп 48
Қой 54 Ноқат 14
Ешкі 60 Арпа мен қара бидай 24
Қоян 44 Жүгері 20
Тауық 78 Кәдімгі зығыр 30
5-сурет. Әртүрлі жануарлар мен өсімдіктердің соматикалық
клеткаларының метафаза кезіндегі кариотиптері.
1-орман бақасынікі (26-хромосома); 2-қойдікі (54 хромосома); 3-адамдікі
(46-хромосома); 4-мысықтың аскаридасынікі (8-хромосома); 5-масанікі (6-
хромосома); 6-дрозофила шыбынынікі (8-хромосома); 7-қара бидайдікі (14-
хромосома); 8-қатты бидайдікі (42-хромосома); 9- сары бөрі бұршақтікі (48-
хромосома); 10-тарынікі (36-хромосома); 11-тауықтікі (78-хромосома); 12-
мүйізді ірі қара малдікі (60-хромосома); 13-жылқынікі (66-хромосома).
17. 17
2-кесте
1-тапсырма. Митоздық бөліну. Кестені толтыру
Фазаның атаулары Фазадағы
хромосомалардың
өзгеруі
Жасушдағы болатын
басқа өзгерістер
3-кесте
2-тапсырма. Кестені толтыру
Салыстыру Ұқсастығы Айырмашылығы Биологиялық маңызы
1. Митоз
2. Мейоз
Қорытынды және есеп беру
1. Тапсырмадаберілген жұмысты орындау, жұмысқаталдау жасау.
2. Тапсырмаларды жұмысдәптеріне түсіру және жұмысты қорғау.
Бақылаусұрақтар:
1. Хромосома деген не? Оны қай уақытта байқауға болады?
2. Хромосома деген атау қайдан шыққан? Олардың көлемi қандай?
3. Морфологиясы бойынша хромосомалар қалай бөлiнедi?
4. Хромосомалардың түрлері мен негізгі қасиеттері?
5. Хромосомалардың гаплойдты және диплойдты жиынтығы дегеніміз не?
2-тақырып. Тұқым қуудың заңдылықтары. Жыныстық көбейудің
негіздері. Менделизм.
Табиғатта тіршілігі мен құрылыс ерекшеліктері сан алуан түрлер тіршілік
етеді. Сол түрлерге жататын жекелеген организмдер жынысты немесе
жыныссыз көбею нәтижесінде өздерінің ерекшеліктерін (морфологиялық,
анатомиялық, физиологиялық, биохимиялық т.б.) тұқым қуалаушылық арқылы
өз ұрпақтарына береді. Тұқым қуалай отырып, ұрпақтан-ұрпаққа берілуі
нәтижесінде өсімдіктер, жануарлар және микроорганизмдердің нақтылы түрге
тән ерекшеліктері мен белгілері сақталады.
Тұқым қуалау заңдылықтарынзерттеу тірі табиғат құрылымыныңәр түрлі
деңгейлерін (молекулалық, хромосомалық, клеткалық, организмдік және
прпуляциялық) қамтиды. Осыған байланысты тұқым қуалауды зерттеуде
18. 18
бірнеше әдістер (гибридологиялық,цитологиялық, популяциялықбиохимиялық
және т.б.) қолданылады.
Тұкым қуалау заңдылықтарын зерттеудің алғашқы негізін салған 1865
жылы ГрегорМендель. Г. Мендель ашқан тұқым қуу заңдылықтары туралы ілім
– менделизм деп аталады. Г. Мендель өз еңбегін өсімдікті будандастырып ұзақ
уақыт тәжірибе жүргізу арқылы дамытты. Оның тәжірибені дұрыс қоя білуі,
тәжірибені жемісті болуына ықпалын тигізді. Ол будандастыру үшін
бұршақтың бір ғана нақты белгісі бар таза сортын алды. Мендель ұзақ уақыт
тәжірибе жасау нәтижесінде бұршақтың 22 түрінен 10 мың будан алды. Ол ең
алғаш тұқым қуалау заңдылықтарын ашты.
Мендельдің бірінші заңы – бірінші ұрпақтардың біркелкілік заңын
көрсетуүшін ғалым асбұршақ өсімдіктерін моногибридтішағылыстыруарқылы
тәжірибе жүргізді. Екі организмнің шағылысуы гибридизация (будандастыру)
деп аталады. Әр түрлі тұқымқуалаушылық материалы бар екі дараны
шағылыстырудан алынған ұрпақтарды гибридті (буданды), ал жеке дараны
гибрид (будан ) деп атайды. Моногибридті шағылыстыру деп альтернативті
(бірін-бірі жоққа шығаратын) белгілердің бір жұбы бойынша бір-бірінен
ерекшеленетін екі организмді шағылыстыруды айтамыз.
Ата-аналық организмдердегі альтернативті жұп белгілердің тұқым
қуалауын оларды будандастыру арқылы зерттейді. Сол жұп белгілердің санына
қарай будандастыру моногибридті, дигибридті және полигибридті деп
ажыратылады.
Альтернативті немесе қарама-қарсы бір жұп белгілері бойынша
ажыратуға болатын ата-аналық формалар будандастырылса, оны моногибридті
деп айтады. Мысалы Мендель тәжирибелері үшін альтернативті жеті белгілері
бойынша ажыратылатын бұршақтың әртүрлі сорттарын тандап алды: тұқымы
сары немесе жасыл, тұқымы тегіс немесе бұдыр, тұқым қабығы сұр немесе ақ,
бойы биік немесе аласа т.б.
Өздігінен тозаңданатын бұршақтың осы сорттарын бір-бірімен
будандастырудың нәтижесінде Мендель тұқым қуалаудың негізгі
заңдылықтарын ашты. Аналық өсімдік ретінде қандай сорттың
пайдаланғанына қарамастан, будандасудан алынған Ғ1 гибридтерінде
альтернативті жұп белгінің тек біреуі ғана көрініс береді. Мұндай белгіні
Мендель доминантты деп атады. Ол жұп белгілердің ішінен: тұқымның
сары түсі жасыл түске, тегістігі тұқым бұдырлығына, тұқым қабығының
сұр түсі ақ түске, бойының биіктігі аласалығына қарағанда доминанттылық
көрсететінін байқады.
Бірінші гибрид ұрпақта көрінбеген белгілерді Мендель рецессивті
белгілер деп атады. Доминантты белгілерді Мендель жоғарыда атап
өткендей латын алфавитінің үлкен әріпімен, рецессивтік белгілерді - кіші
әріпімен белгіледі. Тұқым қуалауға талдау жасағанда бірінші гибрид ұрпақ
бірдей пішінді, бір типтес болғандықтан бұл құбылыс Мендельдің бірінші
заңы немесе бірінші ұрпақ будандарының біркелкілік заңы деп аталады.
Бұл заңды домтанттылық ережесі деп те атайды. Бұл барлық өсімдіктерге,
жануарларға, адамға да тән жалпы құбылыс.
19. 19
Сары Жасыл Тегіс Бұдыр
Сары Тегіс
6-сурет. Мендельдің доминанттылық заңы.
Осы құбылысқа төмендегідей тұжырымдама беруге болады: әр түрлі таза
тармақтарға жататын (гомозиготалы организмдерді) бір-бірінен альтернативті
белгілердің бір жұбы бойынша ерекшеленетін екі организмді шағылыстыру
кезінде алынған бірінші ұрпақтың барлық гибридтері (Ғ1) біркелкі болады және
ата-анасының біреуінің ғана белгісін алып жүреді.
Егер бұршақ өсімдіктердің бірінші гибрид ұрпағы Ғ, өздігінен тозаңданса,
олардың екінші Ғ2 ұрпағында атасының да, анасының да белгілері көрінетін
болады. Мысалы, бұршақтың сары және жасыл тұқым жарнақтары түрлерін
будандастырудан алынған бірінші ұрпақтың түсі сары болады. Ал осы Ғ1
гибридтерін өздігінен тозаңданудан алынған Ғ2 ұрпағында сары жасыл
тұқымдары бар өсімдіктер пайда болады. Яғни бірінші будан ұрпақта
көрінбеген белгілер (жасыл түс) екінші ұрпақта көрінеді. Доминантты және
рецессивті белгілер Ғ2 ұрпағында белгілі бір сандық ара қатынаста болады.
Мендельдің тәжірибесінде Ғ1-де алынған 258 өсімдіктің өздігінен
тозаңдануынан Ғ2-де 8023 тұқым алынды. Осы тұқымдардың 3/4 бөлігі (яғни
6022) сары ал 1/4 бөлігі (яғни 2001) жасыл түсті болып шыққан. Сонымен Ғ2-
алынған доминантты және рецессивті белгілері бар тұқымдардың сандық
қатынасы 3:1 болды. Бір жұп белгілердің осындай арақатынаста ажырауы
Менделъдің екінші заңы немесе ажырау заңы деп аталады.
Жүргізген зерттеулеріне талдау жасаудың негізінде Мендель қорытынды
жасады: рецессивті бастамалар гетерозиготалы организмде жоғалмайды,
өзгеріске ұшырамайды және көбею кезінде дәл сондай рецессивті
бастамалармен кездескен кезде, яғни келесі ұрпақтарда қайтадан көрінеді.
Мысалы, гетерозиготалы организм Аа-дан түзілген А және а гаметалары
бірімен-бірі араласпайтын "таза" гаметалар. Тұқым қуалаушы бастамалардың
будандастыру кезінде бір-бірімен араласпай ұрпақтарына тұрақты берілуі
тұқым қуалаудың дискреттілігін (оқшаулығын) көрсетеді.
Егер доминантты және альтернативті рецессивті белгілерді (мысалы, тегіс
және бұдыр тұқымдар) А және a - әріптері арқылы белгілесек онда Мендель
жасаған тәжірибені схема түрінде былай көрсетуге болады.
Р-мен белгіленген константты ата-ана формаларының әрқайсысында тұқым
қуалау бастамаларының тек бір ғана типі кездеседі - гомозиготалы АА немесе
20. 20
аа. Ондай гомозиготалы организмдер тиісінше А немесе а гаметаларын түзеді.
Осы гаметалардың бірігуіне Ғ1-де гетерозиготалы будан (Aа) ұрпақ пайда
болады. Доминанттылык құбылысына байланысты Ғ1-дегі өсімдіктердің
бәрінің тұқымдарының беті тегіс болып шығады. Ал Ғ2 де А және а
гаметалардың кездейсоқ комбинациялануы әтижесінде белгілердің ажырау
зандылығы байқалады. Яғни, Ғ2 өсімдіктері тұқымдарының 3/4 бөлігінің беті
тегіс (доминантты бастама), 1/4 бөлігінің беті бұдыр (рецессивті бастама)
болып шығады.
7-сурет. Бұршақ тұқымдарының түсі мен пішініне байланысты тұқым
қуалау белгілері, Пиннет торы.
Әр түрлі типті гаметалардың өзара қосылу мүмкіндіктерін есептеп
шығаруды жеңілдету үшін ағылшын генетигі Пеннетұсынған тор жасалынады.
Пеннет торында аталық гаметалар горизонталь бағытта, аналық гаметалар
вертикаль бағытта жазылады. Тордың ішінде гаметалардың комбинациялары
орналастырылады, ол комбинациялар организмдердің генотиптерін көрсетеді.
Мысалы, гомозиготалы АА немесе гетерозиготалы Аа өсімдіктер
тұқымдарының беті тегіс болғанымен олардың тұқым қуалау бастамалары
21. 21
бірдей емес. Организмдердің сыртқы көрінетін белгілерінің жиынтығын
фенотип, ал тұқым қуалау бастамаларының жиынтығын генотип деп атайды.
Пеннет торынан ажыраудың генотип бойынша 1 АА : 2 Аа : 3 аа, ал фенотип
бойынша 3АА : Іаа қатынасында болатындығын оңай көруімізге болады.
Толымсыз доминанттылық. Гетерозиготалы организмде доминантты
ген рецессивті геннің көрінуін үнемі баса алмайды. Бірқатар жағдайларда
бірінші ұрпақ Ғ1 гибриді ата-аналар белгілерінің бірде-бір вариантын толық
көрсетпейді, белгінің көрінуі доминантты немесе рецессивті жаққа көбірек
немесе азырақ ауытқуы байқалатын аралық сипатқа ие болады. Бірақ осы
ұрпақтың барлық даралары бұл белгі бойынша біркелкілік танытады. Мысалы,
қызыл түсті гүлдері бар тансұлуды (намазшам гүлін) (АА), ақ гүлдері бар
өсімдікпен (аа) шағылыстыру нәтижесінде Ғ1 ұрпағында аралық қызғылт түсі
бар гүл (Аа) пайда болады, барлық Ғ1 ұрпақтары біркелкі және барлығының
түсі қызғылт түс болады, ал екінші ұрпақта 25% қызыл түсті гүлдер, 50%
қызғылт түсі бар гүлдер, 25% ақ гүлдері бар өсімдіктер пайда болады.
Толық емес доминанттылық – кең тараған құбылыс. Бұл құбылыс есінек
гүлі түсінің, құс мамығы құбылысының, ірі қара малдың және қой жүнінің түсі,
адамдағы биохимиялық белгілерінің және т.б. белгілердің тұқым қуалауын
зерттеген кезде анықталады.
Толымсыз доминанттылық жағдайында Ғ2 де генотиптер мен
фенотиптердің арақатынасы сәйкес келеді (1АА:2Аа:1аа), ол толық
доминанттылықта гетерозиготалар мен доминантты гомозиготалар бірдей
белгіні көрсетеді. Бұл фенотип бойынша 3:1 қатынасындай болып ажыраудың
өзгеруіне әкеп соғады.
Мендель ашқан белгілердің ажырау заңын хромосомалар ерекшелігімен -
олардың жұп болатындығымен, гомологты хромосомалардың мейоз кезінде
ажырауымен және ұрықтану процесінде олардың қайта қосылып бірігуімен
түсіндіруге болады.
Бұршақ тұқымыныңсары АА және жасыл аа түсі бойынша гомозиготалы
ата-аналық формалар будандастырылды десек олардың бойындағы жұп
аллельдер гомологты хромосомалардың қарама-қарсы локустарында
орналасады. Мейоз нәтижесінде әр гаметада хромосома жұбының тек біреуі
ғана қалады және гомозиготалы организмдегаметалардыңтек бір ғана сорты (А
немесе a) түзіледі. Ұрықтану нәтижесінде гибрид зиготада хромосома жұбы
қайта қалпына келеді, ал Ғ1 гибридінің генотипі А және a гаметаларының
қосылуынан түзілетіндіктен гетерозиготалы диплоидты өсімдіктің генотипі Аа,
ал тұқымның фенотипі сары болады.
Мейоз кезінде ҒI өсімдіктерінің аталығында да және аналығында да жұп
хромосомалар ажырапәр түрлі гаметаларға кетеді, ажырығанхромосомалармен
бірге А және а аллельдері бар аталық және дәл сондай алельдері бар аналық
жыныс клеткалары түзіледі әрі олар тең мөлшерде болады. Ұрықтану кезінде
екі сортты аталық пен екі сортты аналық гаметалардың қосылу мүмкіндігі
бірдей болады.
Талдаушы будандастыру.
Біз қарастырған мысалымызда аналық ретінде рецессивті жасыл
тұқымдары Ір бұршақ өсімдігін алдық. Ал егер доминантты белгі болса, тұқым
22. 22
қуалаудың 3:1 немесе 1:2:1 сипаты өзгеруі мүкін бе? Бұл сұраққа жауап беру
үшін реципрокты будандастыру жүргізу керек. Реципрокты будандастыру
деп доминантты және рецессивті белгісі бар организмдері аналық ретінде де,
аталық ретінде де пайдалануды айтады. Реципрокты будандастыру кезінде туpa
және кері будандастырулар жүргізіледі. Мысалы, АА х аа - тура, ал аа х АА -
кері будандастыру.
Көпшілік жағдайда реципрокты будандастырудан бірдей нәтижелер
алынады, яғни бірінші гибрид ұрпақ Ғ1 біркелкі, сол сияқты Ғ2 кезіндегі
белгілердің ажырауында да ешқандай айырмашылық болмайды. Белгілердің
тұқым қуалауының хромосомалықмеханизмі бұл заңдылықты да оңай түсіндіре
алады.
Белгілердің тұқым қуалауын гибридологиялық анализ әдісімен зерттеуде
кейде қайыра будандастыру қолданылады. Ол бірінші гибрид ұрпақ Ғ1-ді
гомозиготалы (доминантты немесе рецессивті) формалар мен будандастыру
болып табылады және осындай будандастырудан алынған ұрпақты Ғa деп
белгілейді. Оны былайша жазуға болады: Аа х АА немесе Аа х аа. Генетикалық
талдау және селекция жұмыстары үшін осы екі будандастырудың маңызы
бірдей емес. Бірінші гибрид ұрпақ Ғ1-ді доминантты аллельдер бойынша
гомозиготалы формамен қайыра будандастырған кезде оның ұрпақтары
генотипі жағынан 1AA : Іаа болғанымен фенотипі бойынша белгілердің
ажырауы байқалмайды. Ғa кезінде барлық өсімдіктерде тек сары түсті тұқым
пайда болады.
Генетикалық талдау үшін де, сол сияқты селекция жұмыстары үшін де Ғ1
ұрпағын рецессивті аллельдер бойынша гомозиготалы формамен
будандастырудың үлкен маңызы бар. Будандастырудың мұндай жолын
талдаушы (анализдеуші) будандастыру деп атайды.
4-кесте.
Мендельдің тәжірибесі бойынша бұршақ тұқымдарының пішіні мен
түсіне байланысты тұқым қуалау нәтижелері
Бұршақ тұқымының
пішініне мен түсі
Нақты нәтиже Күтілген нәтиже
Тегіс сары 55 52
Тегіс жасыл 51 52
Бұдыр сары 49 52
Бұдыр жасыл 53 52
Барлығы: 208 208
Рецессивті гомозиготалы форма (aа) бір типтес гаметалар (а) түзеді ал
гетерозиготалы гибрид (Аа) екі сортты А және а гаметаларын түзеді. Аналық
жыныс клеткаларының әрқайсысы бірдей мүмкіндікте ұрықтанған
жағдайда Аa және аа генотиптері 1:1 қатынасында болып шығады. Қайыра
будандастырудың осындай жолымен алынған Аа ұрпағы тұқымының сары, аа
ұрпағы тұқымының түсі жасыл, яғни фенотиптері бойынша ұрпақтардың
ажырауы 1:1 қатынасында болады. Сонымен, Ғa кезінде гендердің
ажырау ерекшеліктеріне қарап гибридтің генотипін, гибридтер түзетін
гаметалар типін және олардың арақатынасын талдауға болады. Сондықтан да
23. 23
гибрид организмді рецессивті гомозиготамен будандастыруды
анализдеуші деп айтады. Анализдеуші будандастырудың көмегімен
организмнің белгісіз генотипін анықтауға болады. Мысалы, тұқымның
түсі сары өсімдіктерді жасыл тұқымды өсмдіктермен будандастырған кезде
алынған ұрпақтың тұқымдарының бәрінің түсі сары болса, онда будандастыру
үшін алынған өсімдік тұқымыныңгенотипі АА, яғнидоминантты гомозиготалы
болғаны. Егер дәл осындай будандастырудан алынған сары және жасыл
тұқымды өсімдіктердің арақатынасы 1:1 қатынасында болса, онда анықтауға
алынған сары тұқымды өсімдіктердің генотипі Аа, яғни гетерозиготалы
болғаны. Талдаушы будандастырудың практикалық жұмыста да маңызы зор.
Бақылау сұрақтары
1. Мендель ашқан заңдылықтардың мәні неде?
2. Моногибридті будандастыру кезінде белгілердің ажырауы қалай жүреді?
3. Моногибридті будандастыруларға тән генотиптер мен фенотиптердің
арақатынасы қандай болады?
4. "Генотип" және "фенотип" деген терминдер қандай ұғым береді?
5. Қандай будандастыру талдаушы деп аталады және неге?
№ 2 Практикалық жұмыс
Тақырыбы: Моногибридті шағылыстыру нәтижелерін талдау.
Гибридтердің бірінші ұрпақтарының біркелкілік заңы.
Сабақтың мақсаты: Моногибридті шағылыстыру кезіндегі
тұқымқуалаушылықтың негізгі заңдылықтарымен таныстыру, бірінші ұрпақ
гибридтерінің доминантты және рецессивті белгілерін анықтау әдісін үйрету.
Құрал-жабдықтар: Нұсқау карталары, суреттер, кестелер.
Гибридологиялық талдау нәтижелерін символикалық әріптермен жазу
белгiлерiн негiзiнен Г. Мендель жасады. Оны мысал арқылы көрсетейiк.
Қойдың қарада қара және ақ түстерi кездлеседi. Ондай түстi қойларды
шағылыстырғанда ұрпақтары тек қара түстi болады. Мұндай бiрiн бiрi жойып
жiберетiн белгiлердi альтернативтi белгiлер деп атайды. Альтернативтi
белгiлерге сиырдың мүйiздi немесе тұқыл болуы, тауықтардың роза тәрiздi
немесе жапырақша желбезектерi, қойларда олардың құлақты немесе шұнақ
болуы сияқты белгiлер жатады. Әрбiр алыернативтi белгiлердiң жұбын
анықтайтын аллелдi гендер болады. lрi қараның қара түсiн анықтайтын – қара
түс генi, қызыл түсiн анықтайтын – қызыл түс гендерi болады. Аллелдi гендер
бiр геннiң екi түрлi бейнесi. Демек, қара түс генi мен қызыл түс генi – iрi
қарадағы түстерiнiң екi аллелi, ал тұқылдық пен мүйiздiлiк гендерi де аллелдi
гендер болып табылады. Аллелдi гендерді генетикада тек бiр әрiппен
белгiлейдi. Негiзiнен екiншi бiр белгiге басымдылық көрсететiн доминантты
белгiнiң латынша немесе ағылшынша атының бiрiншi әрпiмен белгiлеу
келiсiлген. Мысалға iрi қараның қара және қызыл түстерiн В және в әрiптерiмен
24. 24
белгiлейдi. Ол “Black” – қара сөзiнен шыққан. Тауықтардың роза және жапырақ
тәрiздi айдарларын гибридтiк талдау кезiнде R және r деп белгiлейдi. Ол
ағылшынша роза сөзiнен шыққан. Кейде аралық тұқым қуу кезiнде, мысалға
андалуз тауықтарының қауырсынының түсiнiң тұқым қууын көрсеткенде қара
қауырсындыларын В, ал оның аллелi ақшыл түстi В1 деп белгiлейдi, В1 – дегенi
олардағы доминантты белгiнiң жоқтығын көрсетедi. Кейде гибридтi талдауды
көрсеткендебұдан да күрделi белгiлеулер кездесiп қалады. Мысалға, қандағы А
гемоглобинiн анықтайтын гендi НвА, ал оның аллелi В гемоглобинiн
анықтайтын гендi НвВ деп белгiлейдi, Нв – латынша гемоглобин сөзiнiң
қысқартылған белгiсi.
Жалпы гибридологиялық талдауды жазудың белгiлi тәртiбi қалыптасқан.
Ата-аналары қатарын Р – әрпiмен белгiлейдi, ол латынның парента - ата-ана
сөзiнен шыққан. Осы әрiптен кейiн будандастырылушы, шағылыстырылушы
немесе қолдан тозаңдандырылушы ата-аналардың генотипi жазылады. Аналық
жынысты генотип ♀ белгiсiмен көрсетiледi (Венераның айнасы деген белгi).
Аталық жынсты генотип - ♂ - белгiсiмен көрсетiледi (Марстың қалқаны мен
найзасы деген белгi). Әдетте ата-аналардың алдымен аналық: одан соң х –
көбейту белгiсi, содан соң барып аталықты көрсетіп жазу қалыптасқан.
Жоғарыдағылар жазылып болған соң төменгi қатарға олар түзетiн
гаметалардың түрлерi жазылады. Одан соңғы қатарға түзiлуi мүмкiн
ұрпақтардыңгенотиптерi жазылады. Ал қатардың басына Ғ - белгiсi қойылады.
Ол “филин” балалар деген сөзден шыққан. Ғ - белгiсiнiң етегiне ұрпақтардың
қатарын бiрiншi ме, екiншi ме, үшіншi ме сан арқылы көрсетедi. (Ғ1, Ғ2, Ғ3).
Гибридтiк талдауды ата-аналар жұбының түзетiн гаметаларынан бастайды.
Моногибридтi будандастыру кезiнде оларды анықтау өте оңай. Бәрiмiзге
белгiлi, жыныс клеткалары жетiлгенде мейоз процесi кезiнде хромосомалар
жұбы, әрине онымен бiрге ондағы аллелдi гендер де екi жыныс клеткасына,
гаметаларға ажырайды. Нәтижесiнде гаметаларға тек бiр ғана аллелдi ген бара
алады. Ендi АА, Аа және аа генотиптерiнен қандай гаметалар түзiлуi мүмкiн
екендiгiн қарастырайық. Гомозиготалы АА генотипiнен мейоз кезiнде тек А
типтi гаметалар түзiлуi мүмкiн, аа генотипiнен де тек бiр типтi а аллелдi
гаметалар түзiлуi мүмкiн. Ал Аа генотипiнен мейоз кезiнде түзiлген
гометолардың 50% А типтi болса, 50% а типтi гаметалар болады. Басқаша
айтқанда гетерозиготалы организмдер гамета түзгенде жартысы доминантты
гендермен (А) болса, жартысы рецессивтi (а) гендермен болады.
1 тапсырма. Бұршақтың тұқым қабығы сұр немесе ақ, бойы биік немесе
аласа белгілерінің қайсысы доминантты. 6-суреттей сызбасын жасаңдар.
2 тапсырма. Құндыздың қалыпты түсi (А) көкшiл, (а) алеут құндызының
түсiне доминантты. Қалыпты құндыздардан мейоз кезiнде қандай гаметалар
типi түзiледi? Олар гомозиготалы болған жағдайда. Ал қалыпты құндыздар
алеут құндыздарының түсiмен гетерозиготалы болған жағдайда, олардан
25. 25
қандай гаметалар типтерi түзiледi? Ол гаметалардың арақатынасы қандай
болады?
3
3 тапсырма. Сиырлардың
шортгорн тұқымында олардың
қызыл түсiн R генi анықтайды, ал ақ
түсiн r генi анықтайды. Ал олардың
гетерозиготаларының түсiнiң түсi
бурыл болады.
а) егерде ақ сиырды қызыл бұқамен
шағылыстырса бiрiншi буын
ұрпақтарының түсi қандай болады?
ә) егерде бурыл сиырды бурыл
бұқамен шағылыстырса бiрiншi
буын ұрпағының түсi қандай
болады?
8-сурет.Ірі қара малдың түсіне байланысты тұқым қуалауы.
4 тапсырма. Мүйiздi қой мен тұқыл қошқарды шағылыстырғанда олардан
қандай ұрпақ алынады?
1. Осы щағылыстыруда еркек қозылар қайсысының қасиетiн, ұрғашы қозылар
қайсысының қасиеттерiн алады?
2. Тұқыл қойлар мен мүйiздi қошқарларды шағылыстырғанда егерде екеуi де
гомозиготалы болса, ұрпақтары қандай болады?
3. Бұл шағылыстыруда еркек қозылар қайсысының қасиетiн, ұрғашы қозылар
қайсысының қасиетiн қабылдайды? Тұқылдылық генi Р, мүйiздiлiк Р1.
5 тапсырма. Аталық организмнiң генотипi Кк. Осындай организмдерде
ұрықтың қандай типтерi түзiлуi мүмкiн? Ондай әртүрлi типтегi ұрықтардың
арақатынасы қандай болады? Олардың арақатынасының мұндай болуы қандай
биологиялық процесске байланысты?
Қорытынды және есеп беру
1. Тапсырмадаберілген жұмысты орындау, жұмысқаталдау жасау.
2. Тапсырмаларды жұмысдәптеріне түсіру және жұмысты қорғау.
Бақылау сұрақтары
1. Мендельдің бірінші заңы бойынша, гибридтердіңбірінші ұрпақтарының
біркелкілігі нені білдіреді?
2. Доминантты және рецессивті белгілер дегеніміз не?
3. Доминантты белгiсi бар гомозиготалы организм қанша типтi гаметалар
түзедi?
4. Гетерозиготалы доминантты белгiсi бар организм қанша типтi гаметалар
түзедi?
5. Рецессивтi белгiсi бар организм қанша типтi гаметалар түзедi?
26. 26
№ 3 Практикалық жұмыс
Тақырыбы: Дигибридті және полигибридті шағылыстыру
нәтижелерін талдау. Белгілердің тәуелсіз ажырау заңы.
Сабақтың мақсаты: Дигибридті және полигибридті шағылыстыру
кезіндегі тұқымқуалаушылықтың негізгі заңдылықтарымен таныстыру,
ұрпақтардың гибридтерінің доминантты және рецессивті белгілерін анықтау
әдісін үйрету.
Құрал-жабдықтар: Нұсқау карталары, суреттер, кестелер.
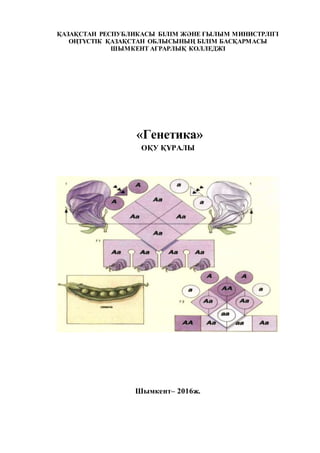
Мендельдің 2-ші заңы - белгілердің тәуелсіз ажырауы. Бұл заң
полигибридтік будандастыруда ашылды. Мұндай будандастыруда бастапқы
формалар екі және одан көп белгілері бойынша айырмашылығы болады.
Мендель ас бұршақтың сары тегіс қабықты сортын (ААВВ) жасыл бұдыр
қабықты сортымен (аавв) будандастырды, яғни олардың альтернативті екі
белгісі - тұқымның түсі және пішіні жағынан айырмашылығы болды.
Будандардың бірінші ұрпағында тұқым сары түсті, оның қабығы тегіс болып
шықты, демек, бастапқы ата-аналық формалар аталған екі белгі бойынша
гомозиготалы болған. Бірінші ұрпақтың генотипі дигетерозиготалы - АаВв.
Өздігінен тозаңданған бірінші ұрпақ өсімдіктерін зерттеп, Мендель F2-де
фенотип бойынша төрт категорияға бөлінген 556 дән алды, атап айтқанда 315-
сары тегіс (АВ), 108-жасыл тегіс (аВ), 101-сары бұдыр (Ав) және 32-жасыл
бұдыр (ав). Мендель 32 санын арақатынас бірлігі ретінде алды, сөйтіп
дигибридтік ажырау формуласын жалпы тұрғыдан алғанда былайша өрнектеді:
9АВ:3Ав:3аВ:1ав.
Дигибридтік ажырасуды талдай отырып, Мендель белгілер жұбының
тәуелсіз ажырауы жөнінде заңдылықты ашты. Дигибридттік будандастырудағы
белгілердің ажырасы, моногибридтік будандаструдағы екі дербес, бір-біріне
тәуелсіз ажыраудың жиынтығы екені көрінеді: 9АВ+3аВ+3Ав+1ав =
/3А+1а/ x /3В+1в/.
Гаметалардың үйлесуін Пеннет торы арқылы көруге болады:
♀ ААВВ х ♂ аавв
сары тегіс бұжыр жасыл
F1 cары тегіс АаВв
5-кесте
F2
АВ Ав аВ ав
АВ ААВВ ААВв АаВВ АаВв
Ав ААВв ААвв АаВв Аавв
аВ АаВВ АаВв ааВВ АаВв
ав АаВв Аавв ааВв Аавв
27. 27
Сонымен, дигибридтік будандастыру кезінде аллельді гендердің екі жұбы
да бір-бірінен тәуелсіз тұқым қуалайды, бұл Мендельдің үшінші заңы. Схемада
көрсетілген генотиптің 16 рет қисындасуы төрт түрлі гаметалардың бір-бірімен
үйлесуінен шыққан. Жұп белгілердің тәуелсіз тұқым қуу комбинативті
өзгергіштіктің негізі болып табылады. Ол өсімдіктер мен жануарларды
будандастырғанда әрдайым байқалады. Сонымен Мендельдің 3-ші заңы
мынандай қорытынды жасауға болады. Әр гомологты хромосомада орналасқан
түрлі гендер бір-біріне тәуелсіз тұқым қуалайды.
Мысалға iрi қараның қара және қызыл түстерiн В және в әрiптерiмен
белгiлейдi. Ол “Black” – қара сөзiнен шыққан. Оны мысал арқылы көрсетейiк.
Ірi қара малда қара және қызыл түстері кездеседi.
♀ ККАА х ♂ аакк
қара түсіті, тұқыл қызыл түсі, мүйiздi
F1 қара түсіті, тұқыл КкАа
F2
9-сурет. Қара түсті тоқал бұқаның, қызыл түсті мүйізді сиырмен шағылысуы
нәтижесі.