Biología VI y VII
•Als PPT, PDF herunterladen•
0 gefällt mir•3,246 views

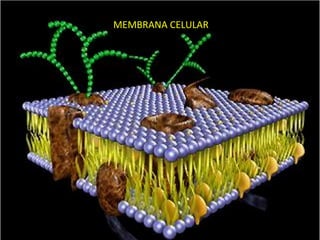
La membrana celular define los límites de la célula y mantiene las diferencias entre su contenido y el entorno. Está compuesta por una bicapa lipídica que actúa como barrera selectiva, permitiendo el paso de nutrientes e impidiendo la fuga de productos celulares. Las proteínas de membrana median sus diversas funciones como el transporte de moléculas, la transducción de señales y la unión estructural. La membrana es crucial para regular las interacciones de la célula con su entorno.

Empfohlen
Weitere ähnliche Inhalte
Was ist angesagt?
Was ist angesagt? (20)
Andere mochten auch
Andere mochten auch (11)
Ähnlich wie Biología VI y VII
Ähnlich wie Biología VI y VII (20)
Kürzlich hochgeladen
Kürzlich hochgeladen (20)
Biología VI y VII
- 2. • La membrana plasmática, que rodea a todas las células, define la extensión de la célula y mantiene las diferencias esenciales entre el contenido de ésta y su entorno. • Esta membrana es algo más que una barrera pasiva. Es también un filtro altamente selectivo que mantiene la desigual concentración de iones a ambos lados de ella, y además permite que los nutrientes penetren y los productos residuales salgan de la célula.
- 3. • Las moléculas protéicas están «disueltas» en la bicapa lipídica y median las diversas funciones de la membrana: algunas sirven para el transporte de moléculas específicas hacia el interior y el exterior de la célula; otras son enzimas que catalizan reacciones asociadas a la membrana; finalmente, otras actúan de eslabones estructurales entre el citoesqueleto de la célula y la matriz extracelular, o de receptores que reciben y traducen las señales químicas procedentes del entorno de la célula.
- 4. • Todas las membranas biológicas, incluidas la membrana plasmática y las membranas internas de la células eucarióticas, tienen una estructura general común: se trata de agrupaciones de moléculas lipídicas y protéicas, unidas por interacciones no covalentes. • Las moléculas lipídicas están dispuestas en forma de una doble capa continua • Esta bicapa lipídica constituye la estructura básica de la membrana y actúa de barrera impermeable al flujo de la mayoría de moléculas hidrosolubles.
- 5. • Las membranas son estructuras asimétricas: la composición lipídica y protéica de sus dos caras se diferencia de manera que refleja las diferentes funciones realizadas por las dos superficies. • Los lípidos de las membranas son moléculas anfipáticas que espontáneamente forman bicapas. • Las moléculas lipídicas son insolubles en agua pero se disuelven fácilmente en disolventes orgánicos. • Constituyen aproximadamente un 50% de la masa de la mayoría de membranas plasmáticas de las células animales. • Existen unas 5 x 106 moléculas lipídicas en una sección de bicapa lipídica de 1nm x 1 nm, o aproximadamente 109 moléculas lipídicas en la membrana plasmática de una célula animal pequeña.
- 6. • Los tres tipos principales de lípidos de las membranas celulares son: los fosfolípidos (los más abundantes), el colesterol y los glucolípidos. • Los tres tipos son anfipáticos, es decir, tienen un extremo hidrofílico (que se siente atraído por el agua o polar), y un extremo hidrofóbico (que rehúye el agua o no polar).
- 7. • Una molécula típica de fosfolípido, tiene una cabeza polar y dos colas hidrocarbonadas hidrofóbicas. • Las colas varían de longitud (normalmente de 14 a 24 átomos de carbono. • Una de ellas contiene generalmente uno o más dobles enlaces cis (es decir, es insaturada) • La otra carece de ellos (es decir, es saturada). • Cada doble enlace cis genera una curvatura en la cola. • Estas diferencias de longitud y grado de saturación entre las colas son importantes porque afectan la fluidez de la membrana.
- 8. • Cuando las moléculas anfipáticas están rodeadas por todas partes por un ambiente acuoso, tienden a agregare escondiendo de dos maneras: pueden formar micelas esféricas, con las colas hacia el interior, y pueden formar láminas bimoleculares , o bicapas.
- 9. En medio acuoso, cada molécula de lípido obliga a las moléculas de agua vecinas a adoptar estados más ordenados (las que están sombreadas de color azul) Cuando las moléculas de lípido se agregan, sólo están más ordenadas las moléculas de agua que están en contacto directo con el agregado. Al ser menos, la entropía aumenta.
- 10. • La formación de la parte lipídica de las membranas biológicas es un proceso de auto-ensamblado. Los compartimentos formados por bicapas lipídicas tienden a cerrarse de nuevo después de haber sido rotos.
- 11. • Además de sus propiedades de auto-ensamblaje y auto-sellado, una bicapa lipídica tiene otras características que la convierten en una estructura ideal para las membranas celulares. Una de las más importantes es la fluidez. • La bicapa lipídica es un fluido bidimensional. • Las moléculas lipídicas de las bicapas artificiales muy rara vez migran de la monocapa de un lado a la del otro lado; este proceso, denominado flip-flop, se produce menos de una vez por semana en cualquier molécula lipídica . • Las moléculas lipídicas intercambian fácilmente su lugar con el de las moléculas vecinas dentro de una monocapa (aprox. 107 por segundo). Esto da lugar a una rápida difusión lateral. • Estudios indican que las moléculas lipídicas giran con gran rapidez alrededor de sus ejes longitudinales y que sus cadenas hidrocarbonadas son flexibles, produciéndose el mayor grado de flexión cerca del centro de la bicapa, y el menor junto a la cabeza polar. • El componente lipídico de una membrana biológica es un líquido bidimensional en el que las moléculas constituyentes se mueven rápidamente, pero generalmente dentro de su propia monocapa.
- 12. – de rotación: es como si girara la molécula en torno a su eje. Es muy frecuente y el responsable en parte de los otros movimientos. – de difusión lateral: las moléculas se difunden de manera lateral dentro de la misma capa. Es el movimiento más frecuente. – flip-flop: es el movimiento de la molécula lipídica de una monocapa a la otra gracias a unas enzimas llamadas flipasas. Es el movimiento menos frecuente, por ser energéticamente más desfavorable. – de flexión: son los movimientos producidos por las colas hidrófobas de los fosfolípidos. • La fluidez es una de las características más importantes de las membranas. Depende de factores como : – la temperatura, la fluidez aumenta al aumentar la temperatura. – la naturaleza de los lípidos, la presencia de lípidos insaturados y de cadena corta favorecen el aumento de fluidez; la presencia de colesterol endurece las membranas, reduciendo su fluidez y permeabilidad. •
- 13. • Otra determinante de la fluidez de una membrana es el colesterol. Las membranas plasmáticas de los eucariotas contienen cantidades relativamente elevadas de colesterol, hasta una proporción de una molécula de colesterol por cada molécula de fosfolípido. • Además de regular la fluidez, se cree que el colesterol aumenta la estabilidad mecánica de la bicapa. • Las moléculas de colesterol se orientan en la bicapa con sus grupos hidroxilo próximos a las cabezas polares de las moléculas de los fosfolípidos; sus anillos esteroides planos interactúan –y en parte inmovilizan– .
- 14. • A las concentraciones en que se presenta el colesterol en la mayoría de membranas plasmáticas de los eucariotas, también ejerce el efecto de impedir que las cadenas hidrocarbonadas se junten y cristalicen. • De esta manera el colesterol inhibe las transiciones de la fase inducidas por la temperatura, impidiendo así el drástico descenso de la fluidez de la membrana que, en caso contrario, se produciría a bajas temperaturas.
- 15. • Las membranas plasmáticas de la mayoría de las células eucarióticas contienen no sólo grandes cantidades de colesterol sino también diversos tipos de fosfolípidos. • La membrana plasmática del eritrocito humano contiene cuatro fosfolípidos principales – fosfatidilcolina, enfingomielina, fosfatidilserina y fosfatidiletanolamina – . • La razón por la que la membrana plasmática de la célula eucariotica contiene una gran variedad de fosfolípidos, es quizá que los lípidos de la membrana actúa como un disolvente bidimensional de las proteínas que necesitan ciertos fosfolípidos específicos para actuar de manera óptima. • La asimetría se genera durante la biosíntesis de la bicapa en el retículo endoplasmático. • Función: ayuda a mantener las proteínas de membrana orientadas adecuadamente en la bicapa.
- 16. • Los glucolípidos se presentan probablemente en las membranas plasmáticas de todas las células animales, donde suelen constituir aproximadamente un 5% de las moléculas lipídicas de la monocapa exterior. • Varían considerablemente de una especie a otra e incluso varían entre los tejidos de una misma especie. • En las bacterias y en las plantas, casi todos los glucolípidos derivan del glicerol, mientras que en las células animales derivan siempre de la esfingosina, un largo amino alcohol.
- 17. • Estos últimos compuestos, denominados glucoesfingolípidos, tienen una estructura general que es similar a las de los fosfolípidos basados en glicerol, ya que poseen un grupo de cabeza polar y dos cadenas hidrocarbonadas hidrofóbicas (aunque una de las cadenas deriva de la esfingosina y la otra es una cadena de ácido graso). Sin embargo, todas las moléculas de glucolípido se distinguen por el hecho de que el grupo de cabeza polar consiste en uno o más residuos de azúcar.
- 18. • Glucolípidos neutros: sus grupos polares de cabeza consisten en un número que oscila entre 1 y 15 o más azúcares neutros. Algunos glucolípidos neutros únicamente se encuentran en ciertos mamíferos. • Un notable ejemplo lo constituye el galactocerebrósido, uno de los glucolípidos más simples, que tan sólo tiene galactosa en su grupo polar de cabeza. Se trata del glucolípido principal de la mielina. • Un rasgo característico de estas células mielinizantes es la gran cantidad de galactocerebrósido que presentan en su membrana plasmática, constituyendo casi el 40% de la monocapa exterior.
- 20. • Los glucolípidos más complejos, los gangliósidos, contienen uno o varios residuos de ácido siálico (conocido también como ácido N-acetilneuramínico o ANAN), que les proporcionan una carga neta negativa. • Los gangliósidos son más abundantes en la membrana plasmática de las neuronas, en donde constituyen aproximadamente un 6% de la masa lipídica total.
- 23. PROTEÍNAS DE MEMBRANA • LA ESTRUCTURA BÁSICA DE LA MEMBRANA ESTÁ DETERMINADA POR LA BICAPA LIPÍDICA, LA MAYOR PARTE DE SUS FUNCIONES ESPECÍFICAS ESTÁN DESEMPEÑADAS POR PROTEÍNAS POR LO TANTO LA CANTIDAD Y TIPO DE PROTEÍNAS DE UNA MEMBRANA REFLEJAN SU FUNCIÓN.
- 24. • Proteínas de membrana • La mayor parte de las funciones específicas de las membranas biológicas están desempeñadas por proteínas. • Muchas proteínas de membrana se mantienen en la bicapa gracias a interacciones hidrofóbicas con las moléculas lipídicas. • El carácter hidrofóbico de algunas proteínas de membrana se ve incrementado por la unión covalente de una o más cadenas de ácidos grasos, que ayudan a anclar estas proteínas en la bicapa.
- 25. • Extracción de la membrana mediante: – Solución salina: Proteínas periféricas – Detergentes o disolventes orgánicos: Proteínas integrales • Muchas proteínas atraviesan la bicapa quedando expuestas a un entorno acuoso en ambos lados de la membrana; reciben el nombre de proteínas transmembrana. • Otras proteínas sólo se hallan expuestas al agua, en un lado de la bicapa; algunas de estas proteínas están ancladas en la membrana mediante interacciones no covalentes con las proteínas transmembrana, mientras que otras pueden estar ancladas mediante la unión covalente a cadenas de ácidos grasos que se encuentran en una monocapa o en la otra.
- 27. • Todas las células eucariotas tienen hidratos de carbono en su superficie en forma de cadenas laterales de oligosacáridos unidos covalentemente a las proteínas de membrana (glucoproteínas) o en menor proporción unidas a los lípidos (glucolípidos)
- 28. • Se cree que la mayoría de las proteínas de membrana que se hallan expuestas en la superficie celular contienen residuos de azúcar, mientras que 1 de cada 2 moléculas de lípidos presenta azúcar • La distribución de los carbohidratos es más asimétrica que la de los fosfolípidos
- 29. • Los términos cubierta celular, glucocáliz y matriz extracelular se utilizan a menudo para describir la zona periférica rica en carbohidratos de la superficie
- 30. • Esquema de la cubierta celular (glucocaliz) que esta formada por las cadenas laterales de oligosacáridos de los glucolípidos y de las glucoproteínas intrínsecas de la membrana y las glucoproteínas y los proteoglicanos absorbidos
- 31. Transporte de moléculas a través de las membranas • La membrana actúa como una barrera altamente impermeable a la mayoría de la moléculas polares, impidiendo que la mayoría del contenido hidrosoluble de la célula salga de ella
- 32. • Por esto mismos la célula ha tenido que desarrollar mecanismos que le permitan: • expulsar de su interior los desechos del metabolismo • adquirir nutrientes del líquido extracelular, • por lo cual se desarrolló la capacidad de la membrana celular de permitir el paso o salida de manera selectiva de algunas sustancias
- 33. • Las membranas celulares permiten el paso del agua y de las moléculas no polares por simple difusión física • Son impermeables a moléculas polares las cuales son transportadas por las proteínas de transporte a través de las membranas
- 34. • CLASIFICACION • Las proteínas poseen veinte aminoácidos, los cuales se clasifican en: Glicina, alamina, valina, leucina, isoleucina, fenil, alanina, triptófano, serina, treonina, tirosina, prolina, hidroxiprolina, metionina, cisteína, cistina, lisina, arginina, histidina, ácido aspártico y ácido glutámico. • Según su composición • pueden clasificarse en proteínas "simples" y proteínas "conjugadas". • Las "simples" o "Holoproteínas" son aquellas que al hidrolizarse producen únicamente aminoácidos • Mientras que las "conjugadas" o "Heteroproteínas" son proteínas que al hidrolizarse producen también, además de los aminoácidos, otros componentes orgánicos o inorgánicos. • La porción no protéica de una proteína conjugada se denomina "grupo prostético". • Las proteínas cojugadas se subclasifican de acuerdo con la naturaleza de sus grupos prostéticos. CONJUGADAS NOMBRE COMPONENTE NO PROTEICO Nucleoproteínas Acidos nucléicos Lipoproteínas Lípidos Fosfoproteínas Grupos fosfato Metaloproteínas Metales Glucoproteínas Monosacáridos
- 35. • Diferentes tipos de movimiento de las moléculas a través de las membranas biológicas. Difusión pasiva y la difusión facilitada no requieren el consumo de energía, ya que se realiza a favor de gradiente de concentración o electroquímico; cuando las sustancias están cargadas la dirección y magnitud del flujo de iones a través de una membrana depende tanto de la diferencia de concentración y de la diferencia eléctrica a través de ella, estas dos fuerzas son por ello colectivamente conocidas como gradiente electroquímico.
- 36. • Algunas sustancias entran directamente en la célula a través de difusión pasiva pero muchas sustancias de interés para la célula atraviesan la membrana mediante difusión facilitada. El transporte activo se realiza con consumo de energía (acoplando a la hidrólisis ATP) al realizarse enana dirección energéticamente desfavorable contra un gradiente electroquímico o de concentración
- 37. • Transporte pasivo- • el paso de una molécula sin carga es determinado por la diferencia de concentración • El paso de una molécula con carga lo va determinar su gradiente de concentración y su gradiente eléctrico total a través de la membrana (potencial de membrana). Ambos gradientes juntos constituyen el gradiente electroquímico
- 38. • Transporte activo- algunas proteínas transportadoras funcionan como bombas que impulsan activamengte el movimiento de los solutos especificos en contra de sus gradientes electroquímicos, se encuentran estrechamente acopladas a una fuente de energía metabólica. Esto implica con frecuencia la hidrólisis de ATP
- 39. • Diferentes tipos de proteínas transportadoras de membrana • Se suelen distinguir dos clases de proteínas que intervienen en la difusión facilitada: • Las proteínas canales o de canal facilitan la difusión formando poros (e.g porinas) abiertos en la membrana que permiten la libre difusión de cualquier molécula de tamaño y carga apropiados. • Las proteínas transportadoras se unen específicamente en un lado de la membrana a las moléculas que van a ser transportadas, sufren entonces un cambio conformacional que permite que la molécula pase a través de la membrana y sea finalmente liberada al otro lado.
- 40. • Se distinguen tres tipos de transportadores: • Uniportes, transportan un solo tipo de molécula a favor de gradiente de concentración • Cotransporte (simportes y antiportes), que catalizan el movimiento de una molécula en contra de su gradiente de concentración dirigido por el cotransporte (en la misma dirección o en sentidos opuestos respectivamente) de otra molécula o ión a favor de gradiente. • Uniporte- transporta un soluto de un lado a otro de la membrana • Cotransporte- la transferencia de un soluto depende de la transferencia simultánea o secuencial de un segundo soluto: • A) simporte- ya sea en la misma dirección • B) antiporte- en la dirección opuesta del transporte de intercambio
- 41. • Existen también canales iónicos (que transportan iones, moléculas cargadas) cuya apertura es regulada por la unión de un pequeño ligando (e.g. un neurotransmisor, acetilcolina) o con una apertura regulada por cambios en el voltaje eléctrico (canales iónicos regulados por voltaje para cationes, etc…).
- 42. • Las bombas potenciadas por ATP permiten a la célula el transporte activo de ciertas moléculas en contra de gradiente de concentración o electroquímico. Los triángulos señalan los gradientes con la punta indicando hacia la zona de menor concentración o de potencial eléctrico.
- 43. • Ejemplos de proteínas que permiten el transporte de iones, denominadas bombas. • El primer ejemplo (más a la derecha) es un complejo de la cadena respiratoria de las mitocondrias. • A continuación una bacteriorodopsina, que usa la luz visible para mover protones a través de la membrana • Por último , una bomba que intercambia sodio y potasio, ayudando a establecer los gradientes de estos iones en la membrana plasmática. (Modificado de Alberts et al., 2002).
- 44. • Proteínas Integrales: Se encuentran ancladas de manera parcial o completa a la bicapa lipidica, las que atraviesan completamente la bicapa pueden tener uno o más de sus dominios anclados a esta. Las hay con un solo cruce como la glicoforina o con varios como algunos receptores. En ambos casos la secuencia o secuencias de aminoácidos localizadas entre las cadenas de ácidos grasos adoptan una conformación en alfa hélice. La aquaporina, un canal que cruza numerosas veces la membrana, posee secuencias de aminoácidos de la zona hidrofóbica que se disponen en hebras beta. (Modificado de Pollard et al., 2007) Alberts, 2002
- 45. • Proteínas periféricas: Se encuentran asociadas a proteínas intengrales, sun función es transitoria Alberts, 2002
- 46. • La gran mayoría de las reacciones metabólicas tienen lugar gracias a la presencia de un catalizador de naturaleza protéica específico para cada reacción. Estos biocatalizadores reciben el nombre de enzimas. La gran mayoría de las proteínas son enzimas. • Las enzimas son muy especificas en el trabajo que realizan. Por ejemplo, las enzimas de amilasa, solo trabajan en almidón, las enzimas de proteasa lo hacen con proteínas, etc., esto permite que las enzimas contengan características que son de gran beneficio celular.
- 48. Vitaminas hidrosolubles , sus coenzimas derivadas y sus funciones Vitamina Coenzima derivada Abreviatura Función Tiamina (B1) Pirofosfato de tiamina TPP Descarboxilación y transferencia de grupos acilo. Riboflavina (B2) Flavina mononucleótido FMN Portadores de hidrógeno y electrones Flavina y adenina FAD en oxido-reducciones dinucleótido Ácido Nicotínico Nicotinamida y adenina dinucleótido NAD+ Portadores de hidrógeno y electrones en oxido-reducciones Nicotinamida y adenina dinucleótido fosfato NADP+ Piridoxina, piridoxal y piridoxamina (B6) Transaminación y decarboxilación Ácido Pantoténico Coenzima A CoASH Transferencia de acilos Biotina Enlazada covalentemente a carboxilasas Carboxilación Ácido Fólico Tetrahidrofolato TH4 Transferencia de un carbono Cobalamina (B12) Coenzima de cobamida Reordenamientos, transferencia de metilos
- 49. Clasificación de las Enzimas (según International Union of Biochemestry) CLASE TIPO DE REACCIÓN CATALIZADA 1 - Oxidorreductasas Síntesis de componentes a tráves de la ruptura oxidativa o reductora de un enlace de alta energía. p. ej. alcohol deshidrogenasa alcohol + NAD+ ® colina + glutamato 2 - Transferasas Transferencia de un grupo funcional de una molécula a otra. p. ej. aspartato aminotransferasa L-aspartato + 2-oxoglutarato ® 2- oxalacetato + 1-L-glutamato 3 - Hidrolasas Ruptura de enlaces por hidrólisis p. ej. Acetilcolina + H2O ® 2 oxalacetato + L- glutamato 4 - Liasas Ruptura de enlaces por eliminación p. ej. Piruvato descarboxilasa 2 oxoácido ® aldehído + CO2 5 - Isomerasas Modificación de la forma o del ordenamiento espacial de las moléculas . p. ej. Fosfoglicerato mutasa 2-fosfoglicerato ® 3- fosfoglicerato 6 - Ligasas Unión de moléculas usando la energía que se deriva de la hidrólisis de los enlaces de alta energía p. ej. Acetil- CoA ligasa ATP + acetato+ CoA ® AMP + acetato + CoA + pirofosfato + acetil-CoA
- 50. • MECANISMO DE ACCIÓN • Como hemos visto las propiedades químicas de una enzima dependen casi enteramente de las cadenas laterales. La actividad catalítica de una enzima resulta de la unión de la molécula de sustrato al sitio activo de la enzima por medio de interacciones generalmente débiles. Al parecer, las más significativas son las interacciones puente hidrógeno. También debemos recordar que esta unión es sumamente específica.
- 51. • Según su función • La diversidad en las funciones de las proteínas en el organismo es quizá la más extensas que se pueda atribuir a una familia de biomoléculas. • Enzimas: Son proteínas cuya función es la "catalisis de las reacciones bioquímicas". Algunas de stas reacciones son muy sencillas; otras requieren de la participación de verdaderos complejos multienzimáticos. El poder catalítico de las enzimas es extraordinario: aumentan la velocidad de una reacción, al menos un millon de veces. • Las enzimas pertenecen al grupo de las proteínas globulares y muchas de ellas son proteínas conjugadas. • Proteínas de transporte: Muchos iones y moléculas específicas son transportados por proteínas específicas. Por ejemplo, la hemoglobina transporta el oxígeno y una porción del gascarbónico desdes y hacia los pulmones, respectivamente. En la memebrana mitocondrial se encuentra una serie de proteínas que trasnportan electrones hasta el oxígeno en el proceso de respiración aeróbica. • Proteínas del movimiento coordinado: El músculo está compuesto por una variedad de proteínas fibrosas. Estas tienen la capacidad de modificar su estructura en relación con cambios en el ambiente electroquímico que las rodea y producir a nivel macro el efecto de una contracción muscular.
- 52. • Proteínas estructurales o de soporte: Las proteínas fibrosas como el colágeno y las a-queratinas constituyen la estructura de muchos tejidos de soporte del organismo, como los tendones y los huesos. • Anticuerpos: Son proteínas altamenmte específicas que tienen la capacidad de identificar susustancias extrañas tale como los virus, las bacterias y las células de otros organismos. • Proteoreceptores: Son proteínas que participan activamente en el proceso de recepción de los impulsos nerviosos como en el caso de la "rodopsina" presente en los bastoncillos de la retina del ojo. • Hormonas y Proteínas represoras: son proteínas que participan en la regulación de procesos metabólicos; las proteínas represoras son elementos importantes dentro del proceso de transmisión de la información genética en la bisíntesis de otras moléculas. •
- 53. • La bomba sodio potasio ATP (adenin-tri-fosfatido) Estructura proteica • es una proteína de membrana que actúa como un transportador de intercambio antiporte (transferencia simultánea de dos solutos en diferentes direcciones) que hidroliza ATP. • Es una ATPasa de transporte tipo P, es decir, sufre fosforilaciones reversibles durante el proceso de transporte. • Está formada por dos subunidades, alfa y beta, que forman un tetrámero integrado en la membrana.
- 54. • La subunidad alfa está compuesta por 10 segmentos transmembrana y en ella se encuentra el centro de unión del ATP que se localiza en el lado citosólico de la membrana. También posee dos centros de unión al potasio extracelulares y tres centros de unión al sodio intracelulares que se encuentran accesibles para los iones en función de si la proteína está fosforilada. • La subunidad beta contiene una sola región helicoidal transmembrana y no parece ser esencial para el transporte ni para la actividad ATPasa. La enzima está glucosilada en la cara externa (como la mayoría de proteínas de membrana) y requiere de magnesio como cofactor para su funcionamiento ya que es una ATPasa.
- 55. • La base iónica del potencial de membrana • 1 está determinado en gran parte por el gradiente de concentración de K+ • 2 la membrana plasmática en reposo es más permeable al k+ que al Na+ o a los aniones • El gradiente de K+ y las diferencias de permeabilidad iónica están determinados, a su vez por las propiedades de las proteínas de transporte específicas de la propia membrana plasmática
- 56. • El transporte activo puede ser impulsado por los gradientes iónicos • Muchos sistemas de transporte activo son impulsados por la energía almacenada en los gradientes iónicos y no directamente por la hidrólisis del ATP todos ellos funcionan como sistemas de cotransporte- algunos como cotransportes unidireccionales otros como transportes de intercambio
- 57. Canales iónicos algunos canales protéicos transmembrana constituyen una puerta que solo se abre transitoriamente pues se encuentran regulados, -se abren en respuesta a un ligando extracelular que se une a un receptor específico de la superficie celular y reciben el nombre de canales regulados por ligando -otros se abren en respuesta a cambios en la concentración intracelular de iones determinados
- 58. • La mayoría de los canales no permanecen abiertos permanentemente, sino que se abren en respuesta a estímulos. Estos estímulos pueden ser tanto la presencia de una sustancia inductora como una modificación de la carga eléctrica de la membrana (modificación del potencial eléctrico). Los canales que se abren o cierran en presencia de sustancias inductoras (ligandos) son llamados dependientes de ligando y los otros, dependientes de voltaje.
- 59. • Carriers (acarreadores) o permeasas: • Al igual que los canales iónicos, las permeasas están formadas por proteínas transmembrana multipaso. Suelen transportar una gran variedad de iones como el HCO3- y otras moléculas polares sin carga como la glucosa. • Este tipo de proteínas fijan una única molécula de sustrato (o unas pocas) a la vez, y a continuación sufren un cambio conformacional reversible que les permite transportar el soluto de un lado al otro de la membrana (translocación). • Para entender la difusión facilitada no hay que pensar si una sustancia “entra o sale” de la célula, lo importante es considerar que se está movilizando algo a favor del gradiente (químico o eléctrico) gracias a la acción de proteínas transportadoras. • Por esta razón es que no se requiere de energía adicional, no se requiere gasto de ATP, ya que es el propio gradiente el que impulsa el pasaje a través de los transportadores. • Este tipo de transporte es siempre sin gasto de energía y a favor del gradiente electroquímico
- 60. Funciones ddee llaass pprrootteeíínnaass ddee mmeemmbbrraannaa
- 62. Concentracion yy ggrraaddiieenntteess eelleeccttrrooqquuíímmiiccooss Dos proteínas son cruciales para generar y mantener el potencial de membrana, la primera es la bomba de sodio-potasio que bombea activamente sodio hacia el exterior de la célula y potasio al interior. La otra es una canal de fuga de potasio que permite que el potasio salga de la célula a favor de su gradiente de concentración
- 63. Equilibrios iónicos y potenciales de reposo La mayoría de las células animales presentan una diferencia de potencial eléctrico (voltaje) a través de sus membranas
- 64. Génesis del potencial de membrana • La distribución de iones a través de la membrana celular y la naturaleza de la membrana nos dan la explicación del potencial de membrana • Como consecuencia de la salida neta de potasio el interior de la célula pasa a ser eléctricamente negativo en relación con el exterior • el potencial de membrana resultante retarda el movimiento hacia afuera del potasio a través del canal de fuga • La tendencia del potasio a salir a causa de su gradiente de concentración se halla exactamente equilibrada por la tendencia de potasio a penetrar en la célula a causa de su gradiente eléctrico (potencial de membrana)
- 65. Fuerzas que actúan sobre los iones • Gradiente de concentración [] • Gradiente eléctrico +- • Se alcanza el equilibrio cuando la entrada y la salida del ión son iguales • El potencial de membrana en el cual se alcanza este equilibrio es el potencial de equilibrio
- 66. La bomba de sodio potasio es electrogénica, porque extrae tres átomos de sodio por cada dos de potasio que introduce por tanto contribuye al potencial de membrana K+ CONCENTRACIÓN K+ ELÉCTRICO INTERIOR DE LA CÉLULA - EQUILIBRIO Exceso de Cationes en exterior ATP-asa Na+-K+ 2K+ 3Na+
- 67. Normalmente el citoplasma suele ser eléctricamente negativo en relación con el líquido extracelular • La diferencia de potencial eléctrico a través de la membrana en condiciones de reposo recibe el nombre de potencial de reposo de la membrana y desempeña un papel muy importante en la excitabilidad de las células nerviosas y fibras musculares, así como en otras respuestas celulares
- 69. • Todas las células capaces de producir potenciales de acción presentan potenciales de membrana de reposo de cuantía considerable • También la mayoría de las células no excitables tienen potenciales de reposo
- 70. • El potencial de reposo es necesario para que la célula pueda disparar un potencial de acción • La bomba Na+ - K+ ATPasa crea gradientes de estos iones a través de las membranas plasmáticas de las células, transfiere una carga neta a través de la membrana y contribuye directamente al potencial de reposo
- 72. • En química, la bomba sodio-potasio es una proteína de membrana fundamental en la fisiología de las células que se encuentra en todas nuestras membranas celulares. Su función es el transporte de los iones inorgánicos más importantes en biología (el sodio y el potasio) entre el medio extracelular y el citoplasma, proceso fundamental en todo el reino animal. • La ATPasa Na+/K+, genera un gradiente de concentración de K+, y es el movimiento de este ión, a través de los canales de K+ de reposo, desde el citosol al exterior celular a favor de su gradiente de concentración el que genera el potencial de membrana negativo en el interior. • Por tanto, son los canales de K+ de reposo, que están siempre abiertos, los que generan el potencial de reposo
- 74. IIoonniicc BBaassiiss ooff RRMMPP
- 75. • Estructura proteica • La bomba sodio potasio ATP (adenin-tri-fosfatido) es una proteína de membrana que actúa como un transportador de intercambio antiporte (transferencia simultánea de dos solutos en diferentes direcciones) que hidroliza ATP. • Es una ATPasa de transporte tipo P, es decir, sufre fosforilaciones reversibles durante el proceso de transporte. • Está formada por dos subunidades, alfa y beta, que forman un tetrámero integrado en la membrana.
- 76. • La subunidad alfa está compuesta por 10 segmentos transmembrana y en ella se encuentra el centro de unión del ATP que se localiza en el lado citosólico de la membrana. También posee dos centros de unión al potasio extracelulares y tres centros de unión al sodio intracelulares que se encuentran accesibles para los iones en función de si la proteína está fosforilada. • La subunidad beta contiene una sola región helicoidal transmembrana y no parece ser esencial para el transporte ni para la actividad ATPasa. La enzima está glucosilada en la cara externa (como la mayoría de proteínas de membrana) y requiere de magnesio como cofactor para su funcionamiento ya que es una ATPasa.
- 77. • Mantenimiento de la osmolaridad y del volumen celular • La bomba de Na+/K+ juega un papel muy importante en el mantenimiento del volumen celular entre el interior y el exterior de la célula existen diferentes niveles de concentración, siendo mayor la concentración de solutos dentro que fuera de la célula. • La bomba extrae de la célula más moléculas de las que introduce tiende a igualar las concentraciones y, consecuentemente, la presión osmótica.
- 78. • Sin la existencia de la bomba, dado que los solutos orgánicos intracelulares, a pesar de contribuir en sí mismos poco a la presión osmótica total, tienen una gran cantidad de solutos inorgánicos asociados, la concentración intracelular de estos (que generalmente son iones) es mayor que la extracelular, se produciría un proceso osmótico, consistente en el paso de agua a través de la membrana plasmática hacia el interior de la célula, que aumentaría de volumen y diluiría sus componentes. • Las consecuencias serían catastróficas ya que la célula podría llegar a reventar (proceso conocido como lisis).
- 79. • Osmolaridad • Es el número de osmoles por litro de solución • Osmolalidad • Es el número de osmoles por kilogramo de solvente
- 81. • La osmolaridad se modifica por el volumen de los diversos solutos en la solución y por la temperatura, la osmolalidad no.
- 82. • Las sustancias con actividad osmótica en el cuerpo se disuelven en agua cuya densidad es 1, así las concentraciones osmolales pueden expresarse como osmoles por litro (Osm/L) de agua
- 83. Concentración osmolal del plasma • Tonicidad se emplea para describir la osmolalidad de un soluto en relación con el plasma • Isotónicas solución que tiene la misma osmolalidad que el plasma • Hipertónicas las que tienen una osmolalidad mayor que el plasma • Hipotónicas osmolalidad menor que el plasma
- 85. • Osmolalidad del plasma 290 mOsm de cada litro de plasma normal la actividad osmótica se debe al Na+ y a sus aniones acompañantes, en particular Cl- y HCO3-
- 86. • La osmolalidad plasmática total es importante para valorar la deshidratación o sobrehidratación y otras alteraciones electrolíticas • La hiperosmolalidad puede causar coma (coma hiperosmolar)
- 87. • Las células eucariotas mantienen concentraciones muy bajas de Ca++ en el citosol, el gradiente se mantiene por las bombas de Ca++ que están en las membranas y que transportan activamente CA++ • En el músculo la bomba de CA++ es responsable del bombeo de Ca++ desde el citosol hacia el retículo sarcoplásmico Canal de calcio del retículo sarcoplasmático canal inactivado citoplasma Membrana del Retículo sarcoplámico lumen inactivación
- 88. • El transporte activo puede ser impulsado por los gradientes iónicos • Muchos sistemas de transporte activo son impulsados por la energía almacenada en los gradientes iónicos y no directamente por la hidrólisis del ATP todos ellos funcionan como sistemas de cotransporte- algunos como cotransportes unidireccionales otros como transportes de intercambio
- 89. Canales iónicos algunos canales protéicos transmembrana constituyen una puerta que solo se abre transitoriamente pues se encuentran regulados, -se abren en respuesta a un ligando extracelular que se une a un receptor específico de la superficie celular y reciben el nombre de canales regulados por ligando -otros se abren en respuesta a cambios en la concentración intracelular de iones determinados
- 90. Secuencia de fenómenos en la contracción del músculo esquelético • 1. descarga de la neurona motora • 2. liberación del neurotransmisor • 3. unión de la Ach con los receptores nicotínicos para acetilcolina • 4. aumento de la conductancia al sodio y al potasio en la membrana de la placa terminal • 5. generación del potencial de placa terminal • 6. generación del potencial de acción en las fibras musculares • 7. propagación de la despolarización hacia el interior a lo largo de los túbulos T • 8. liberación de Ca2+ de las cisternas terminales y difusión de este ión a los filamentos gruesos y delgados • 9. unión del Ca 2+ con la troponina C y se descubren los sitios de unión de la actina con la miosina • 10. formación de enlaces cruzados entre la actina y la miosina y deslizamiento de los filamentos delgados sobre los gruesos, lo que produce el acortamiento
- 91. Exitación
- 93. Mecanismo de deslizamiento de los filamentos en la contracción
- 94. Secuencia de fenómenos en la relajación del músculo esquelético • 1. el calcio se bombea al retículo sarcoplásmico • 2. liberación de calcio de la troponina • 3. cesa la interacción entre la actina y la miosina
- 95. Relaxation of Muscle Fiber
- 96. Secuencia de fenómenos en la contracción del músculo esquelético • 1. descarga de la neurona motora • 2. liberación del neurotransmisor • 3. unión de la Ach con los receptores nicotínicos para acetilcolina • 4. aumento de la conductancia al sodio y al potasio en la membrana de la placa terminal • 5. generación del potencial de placa terminal • 6. generación del potencial de acción en las fibras musculares • 7. propagación de la despolarización hacia el interior a lo largo de los túbulos T • 8. liberación de Ca2+ de las cisternas terminales y difusión de este ión a los filamentos gruesos y delgados • 9. unión del Ca 2+ con la troponina C y se descubren los sitios de unión de la actina con la miosina • 10. formación de enlaces cruzados entre la actina y la miosina y deslizamiento de los filamentos delgados sobre los gruesos, lo que produce el acortamiento
- 99. • La mayoría de los canales no permanecen abiertos permanentemente, sino que se abren en respuesta a estímulos. Estos estímulos pueden ser tanto la presencia de una sustancia inductora como una modificación de la carga eléctrica de la membrana (modificación del potencial eléctrico). Los canales que se abren o cierran en presencia de sustancias inductoras (ligandos) son llamados dependientes de ligando y los otros, dependientes de voltaje.
- 100. • Carriers (acarreadores) o permeasas: • Al igual que los canales iónicos, las permeasas están formadas por proteínas transmembrana multipaso. Suelen transportar una gran variedad de iones como el HCO3- y otras moléculas polares sin carga como la glucosa. • Este tipo de proteínas fijan una única molécula de sustrato (o unas pocas) a la vez, y a continuación sufren un cambio conformacional reversible que les permite transportar el soluto de un lado al otro de la membrana (translocación). • Para entender la difusión facilitada no hay que pensar si una sustancia “entra o sale” de la célula, lo importante es considerar que se está movilizando algo a favor del gradiente (químico o eléctrico) gracias a la acción de proteínas transportadoras. • Por esta razón es que no se requiere de energía adicional, no se requiere gasto de ATP, ya que es el propio gradiente el que impulsa el pasaje a través de los transportadores. • Este tipo de transporte es siempre sin gasto de energía y a favor del gradiente electroquímico
- 101. Funciones ddee llaass pprrootteeíínnaass ddee mmeemmbbrraannaa
- 103. Organización del cuerpo • Compartimientos líquidos • En el humano adulto promedio el 18% del peso corporal lo constituyen las proteínas y sustancias relacionadas, el 7% son minerales y el 15% es grasa; el restante 60% es agua
- 104. Cerca del 60% del cuerpo humano adulto es líquido, la mayor parte de éste líquido se encuentra dentro de las células y se denomina líquido intracelular, casi un tercio se encuentra alrededor de las células y se denomina líquido extracelular
- 105. LEC • En los animales dotados de sistema vascular cerrado el LEC se divide en dos componentes • Líquido intersticial • Plasma sanguíneo circulante
- 106. • El plasma más elementos celulares sanguíneos conforman el volumen sanguíneo total • El liquido intersticial es la parte externa del sistema vascular que baña a las células
- 107. Unidades para medir la concentración de solutos • Cuando se consideran los efectos de varias sustancias fisiológicas y las interacciones entre éstas, el número de moléculas cargas eléctricas, o las partículas de sustancia por unidad de volumen de un líquido corporal en particular son más importantes que el mero peso de la sustancia por unidad de volumen
- 108. • Por esta razón a menudo las concentraciones se expresan en moles, equivalentes u osmoles • Moles • Un mol es el peso molecular-gramo de una sustancia o el peso molecular de una sustancia en gramos
- 109. • Mol incluye 6x10 23 moléculas • mmol y mmol • así • Un mol de NaCl = 23+35.5=58.5g • El mol es la unidad básica para expresar la cantidad de sustancia según el sistema internacional (SI) de unidades
- 110. • Equivalentes • Muchos de los solutos importantes del cuerpo se encuentran en forma de partículas cargadas • 1 equivalente es un mol de sustancia ionizada, dividido entre su equivalencia.
- 111. • Un mol de NaCl se disocia en 1 Eq de Na+ y 1 eq de Cl- • 1 eq de Na+ =23g/1, pero 1 Eq de Ca2+ =40g/2 =20g • La equivalencia eléctrica no siempre es la misma que la equivalencia química
- 112. • Los mecanismos que permiten a las sustancias cruzar las membranas plasmáticas son esenciales para la vida y la comunicación de las células. Para ello, la célula dispone de dos procesos: Transporte pasivo: cuando no se requiere energía para que la sustancia cruce la membrana plasmática • Transporte activo: cuando la célula utiliza ATP como fuente de energía pasa hacer atravesar la membrana a una sustancia en particular • TRANSPORTE PASIVO • Los mecanismos de transporte pasivo son: • Difusión simple • Osmosis • Ultrafiltración • Difusión facilitada
- 113. • La difusión es el proceso por el cual un gas o una sustancia en solución se dispersa por el movimiento de sus partículas para llenar todo el volumen disponible • Las moléculas en solución están dotadas de energía cinética y, por tanto tienen movimientos que se realizan al azar. • La difusión consiste en la mezcla de estas moléculas debido a su energía cinética cuando existe un gradiente de concentración, es decir cuando en una parte de la solución la concentración de las moléculas es más elevada. • La difusión tiene lugar hasta que la concentración se iguala en todas las partes y será tanto más rápida cuanto mayor sea energía cinética (que depende de la temperatura) y el gradiente de concentración y cuanto menor sea el tamaño de las moléculas. Difusión
- 114. • Las partículas (moléculas o átomos) de una sustancia disuelta en un solvente están en movimiento aleatorio continuo. • Existe un flujo neto de partículas del soluto de las áreas de alta a las de baja concentración • Algunas sustancias como el agua, el oxígeno, dióxido de carbono, esteroides, vitaminas liposolubles, urea, glicerina, alcoholes de pequeño peso molecular atraviesan la membrana celular por difusión, disolviendose en la capa de fosfolípidos. • Algunas sustancias iónicas también pueden cruzar la membrana plasmática por difusión, pero empleando los canales constituídos por proteínas integrales llenas de agua. Algunos ejemplos notables son el Na+, K+, HCO3, Ca++, etc. Debido al pequeño tamaño de los canales, la difusión a través de estos es mucho más lenta que a través de la bicapa fosfolipídica
- 115. • La difusión de las moléculas de solvente hacia la región en la que hay mayor concentración de soluto al cual la membrana es semipermeable se llama ósmosis • Es otro proceso de transporte pasivo, mediante el cual, un disolvente - el agua en el caso de los sistemas biológicos - pasa selectivamente a través de una membrana semi-permeable. • La membrana de las células es una membrana semi-permeable ya que permite el paso del agua por difusión pero no la de iones y otros materiales. • Si la concentración de agua es mayor (o lo que es lo mismo la concentración de solutos menor) de un lado de la membrana es mayor que la del otro lado, existe una tendencia a que el agua pase al lado donde su concentración es menor. ósmosis
- 116. • La tendencia del movimiento de las moléculas de solvente hacia la región con mayor concentración de soluto puede prevenirse si se aplica presión a la solución más concentrada • El movimiento del agua a través de la membrana semi-permeable genera un presión hidrostática llamada presión osmótica. La presión osmótica es la presión necesaria para prevenir el movimiento neto del agua a través de una membrana semi-permeable que separa dos soluciones de diferentes concentraciones.
- 117. TTrraannssppoorrttee ddee mmeemmbbrraannaa -- ÓÓssmmoossiiss
- 118. – La ósmosis puede entenderse muy bien considerando el efecto de las diferentes concentraciones de agua sobre la forma de las células. – Para mantener la forma de un célula, por ejemplo un hematíe, esta debe estar rodeada de una solución isotónica, lo que quiere decir que la concentración de agua de esta solución es la misma que la del interior de la célula. En condiciones normales, el suero salino normal (0.9% de NaCl) es isotónico para los hematíes. Si los hematíes son llevados a una solución que contenga menos sales (se dice que la solución es hipotónica), dado que la membrana celular es semi-permeable, sólo el agua puede atravesarla. Al ser la concentración de agua mayor en la solución hipotónica, el agua entra en el hematíe con lo que este se hincha, pudiendo eventualmente estallar (este fenómeno se conoce con el nombre de hemolisis. Por el contrario, si los hematíes se llevan a una solución hipertónica (con una concentración de sales superior a la del hematíe) parte del agua de este pasará a la solución produciéndose el fenómeno de crenación y quedando los hematiés como "arrugados".
- 120. • Transporte de membrana de macromoléculas y partículas : exocitosis y endocitosis • La endocitosis consiste en un invaginación de una región de la membrana plasmática que posteriormente se estrangula para dar lugar a una vesícula intracelular. • De este modo, la célula incorpora, englobándolas en el interior de esta vesícula, partículas procedentes del medio extracelular . • La deformación de la membrana plasmática que acompaña a los procesos de endocitosis está controlada por una proteína del citosol, denominada clatrina, que se polimeriza para formar un revestimiento de aspecto reticular, el cual atrae hacia su interior a la porción de membrana que dará lugar a la vesícula. El revestimiento de clatrina se elimina una vez dicha vesícula se separa de la membrana por estrangulamiento.
- 121. • Un rasgo importante de la endocitosis y exocitosis es que en las macromoléculas segregadas o ingeridas se hallan secuestradas vesículas y no suelen mezclarse con otras macromoléculas o con los orgánulos de las células • Mediante mecanismos desconocidos las vesículas se fusionan solo con la estructura específica de la membrana, asegurando una transferencia correcta de macromoléculas entre el interior y el exterior
- 122. TTrraannssppoorrttee vveessiiccuullaarr ppiinnoocciittoossiiss • Existen dos modalidades de endocitosis en función del tamaño de las partículas incorporadas: • a) Pinocitosis.- Consiste en la incorporación, en forma de pequeñas vesículas, de partículas de pequeño tamaño (incluidas las macromoléculas) que se encuentran en disolución. • La inmensa mayoría de las células tienen la capacidad de incorporar por este procedimiento distintos tipos de sustancias.
- 123. • Algunos procesos de pinocitosis están mediados por receptores específicos de naturaleza protéica que se encuentran en la membrana celular; estos receptores fijan de manera específica sobre la membrana a determinadas macromoléculas, que a continuación son incorporadas en forma de vesículas pinocíticas; de este modo la célula puede incorporar macromoléculas específicas que se encuentran en pequeñas cantidades en el medio extracelular sin tener que acompañarlas de una gran cantidad de líquido. • La pinocitosis mediada por receptores específicos es el procedimiento por el que penetran en la célula determinadas hormonas, el colesterol, e incluso virus y algunas toxinas de origen bacteriano.
- 124. VVeessiiccuullaarr TTrraannssppoorrtt RReecceeppttoorr--MMeeddiiaatteedd EEnnddooccyyttoossiiss • b) Fagocitosis.- Consiste en la incorporación, en forma de grandes vesículas denominadas fagosomas, de partículas de tamaño superior al de las macromoléculas, que se encuentran en suspensión en el medio extracelular. • De este modo, la célula puede incorporar una gran variedad de partículas de tamaño variado: complejos supramoleculares u orgánulos procedentes de células muertas en descomposición, e incluso células enteras. Sólo algunos tipos celulares tienen la capacidad de fagocitar; entre ellos destacan algunos protozoos que se alimentan de partículas orgánicas en suspensión y algunos leucocitos que fagocitan a los microorganismos invasores con el objeto de eliminarlos.
- 126. • La exocitosis es un proceso inverso al de endocitosis, en el que una vesícula intracelular se aproxima a la membrana plasmática fundiéndose con ella de manera que el contenido de dicha vesícula es vertido al medio extracelular .Por exocitosis la célula puede expulsar los restos del proceso de digestión celular que no le resultan útiles y también los productos de secreción procedentes del aparato de Golgi en forma de vesículas secretoras
- 127. • Las proteínas que serán segregadas se sintetizan en los ribosomas del RER, pasan al interior del lumen delRE y son transportadas al complejo de Golgi en vesículas de transporte acá las proteínas son modificadas , concentradas y seleccionadas y finalmente empaquetadas en una vesícula que se separa del CG y se fusionan con la membrana plasmática.
- 128. • Las moléculas que serán segregadas se transportan activamente al interior de la vesícula secretora • Este proceso es impulsado por un gradiente iónico • Las vesículas secretoras se liberan únicamente cuando se estimula la célula por una señal extracelular • La señal generalmente es un mensajero químico como una hormona • Esta señal puede producir un aumento transitorio en la entrada de Ca++ • El aumento de Ca++ inicia la exocitosis liberando su contenido al exterior
- 129. Fuerzas que actúan sobre los iones • Gradiente de concentración [] • Gradiente eléctrico +- • Se alcanza el equilibrio cuando la entrada y la salida del ión son iguales • El potencial de membrana en el cual se alcanza este equilibrio es el potencial de equilibrio
- 130. Concentration && EElleeccttrroocchheemmiiccaall GGrraaddiieennttss
- 131. Génesis del potencial de membrana • La distribución de iones a través de la membrana celular y la naturaleza de la membrana nos dan la explicación del potencial de membrana
- 132. La bomba de sodio potasio es electrogénica, porque extrae tres átomos de sodio por cada dos de potasio que introduce por tanto contribuye al potencial de membrana K+ CONCENTRACIÓN K+ ELÉCTRICO INTERIOR DE LA CÉLULA - EQUILIBRIO Exceso de Cationes en exterior ATP-asa Na+-K+ 2K+ 3Na+
- 133. Potencial de acción generación y conducción El potencial de acción es un cambio rápido del potencial de membrana seguido de un retorno al potencial de reposo
- 134. • La comunicación entre las células nerviosas se basa en una alteración eléctrica que se propaga a lo largo de la membrana y que recibe el nombre de potencial de acción
- 135. • El tamaño y la forma del potencial de acción difiere considerablemente de un tejido excitable a otro • Se propaga con la misma forma y tamaño a lo largo de toda la fibra muscular o célula nerviosa
- 137. El potencial de acción es la base de la capacidad de las células nerviosas para transportar una señal y hace posible la contracción casi simultánea de una fibra muscular
- 138. El potencial de acción se dispara cuando la despolarización es suficiente para que el potencial de membrana alcance un determinado valor umbral
- 139. • El potencial de acción difiere de la respuesta depolarizante local en tres aspectos: • 1) Se trata de una respuesta mucho mayor en la que se invierten la polaridad de la membrana • 2) El potencial de acción se propaga sin decremento a lo largo de toda la fibra nerviosa o muscular • 3) El tamaño y al forma del potencial de acción se mantienen cosntantes a lo largo de la fibra, su magnitud no decrece con la distancia
- 140. PPootteenncciiaall llooccaall Chemical Synapses
- 141. • Cuando se aplica un estímulo de valor umbral la forma y el tamaño del potencial de acción no se modifica; la magnitud no aumenta al aumentar la intensidad del estímulo
- 142. • Un estímulo subumbral no produce un potencial de acción un estímulo umbral produce un potencial de acción • El potencial de acción constituye una respuesta todo o nada
- 144. Mecaninsmos iónicos responsables del potencial de acción • 1.- aumento rápido de la conductancia a Na+ en la fase temprana del potencial de acción • 2.- el pico del potencial de acción no alcanza más que +50mV • 3.- aumento de la conductancia al K+ más lentamente • 4.- el K+ proporciona una tendencia opuesta • 5.- la conductancia del Na+ decrece muy rápidamente hacia su valor de reposo • 6.- retorno del potencial de membrana al potencial de reposo debido a la rápida disminución de la conductancia al Na+ y al aumento de la conductancia al K+
- 145. Canales iónicos y compuertas • Las corrientes iónicas atraviesan las membranas plasmáticas por canales distintos para sodio y potasio cada uno con características propias, para penetrar en el canal deben perder la mayor parte de su agua y pasar a través del filtro de selectividad Canal de sodio
- 146. • El bloqueador específico para los canales de sodio sensibles a TTX (tetrodotoxina) se une al lado extracelular • El canal parece tener una compuerta de inactivación responsable de los cambios de conductancia
- 147. • El tetratetilamonio (TEA) es un bloqueador de los canales de potasio bloquea al canal por la parte interior de la membrana
- 148. • El sistema nervioso central contiene alrededor de 100 000 millones de neuronas, también contiene de 10 a 50 veces éste número de células gliales • Se calcula que el 40% de los genes participan en su formación NEURONAS CELULAS GLIALES
- 150. • Los axones de muchas neuronas están mielinizados, cuentan con vaina de mielina, la mielina es un complejo lipoprotéico que se envuelve alrededor del axón • Fuera del SNC la mielina se produce en las células de Schwan, células similares a las gliales • La vaina de mielina envuelve todo el axón excepto en sus extremos y en los nodos de Ranvier
- 151. • Desde el punto de vista funcional la neurona casi siempre tiene cuatro zonas importantes: • 1.- una zona receptora dendrítica en donde se integran los múltiples cambios del potencial local mediante las conexiones sinápticas • 2.- un sitio en el cual se generan los potenciales de acción que se propagan • 3.- Un proceso axónico que transmite los impulsos propagados a las terminales nerviosas • 4.- las terminales nerviosas donde los potenciales de acción inducen la liberación del transmisor sináptico
- 152. • 1. DENDRITA. Filamento que emerge de la neurona y que transporta la señal eléctrica producida por un estímulo nervioso. 2. TRANSMISOR. El impulso nervioso se traduce en la liberación de una proteína neurotransmisora. 3. IMPULSO NERVIOSO. La información llevada por el neurotransmisor es transformada de nuevo en un mensaje eléctrico. 4. SINAPSIS. El punto de comunicación entre dos neuronas. 5. NEURONA PRESINÁPTICA. 6. AXÓN. Filamento que recibe información de la neurona postsináptica.
- 153. Excitación y conducción • Las células nerviosas tienen un umbral de excitación bajo • El estímulo puede ser químico eléctrico o mecánico • Se producen dos tipos de cambios fisicoquímicos: • 1.- Potenciales sinápticos, generadores o electrotónicos: • producidos por estímulos inferiores al umbral, que tienen un efecto sobre el potencial de membrana • 2.- Potenciales de acción
- 157. SSiiggnnaall CCoonndduuccttiioonn UUnnmmyyeelliinnaatteedd FFiibbeerr CCoonnttiinnuuoouuss CCoonndduuccttiioonn
- 158. SSiiggnnaall CCoonndduuccttiioonn MMyyeelliinnaatteedd FFiibbeerr SSaallttaattoorryy CCoonndduuccttiioonn
- 159. Conducción ortodrómica y antidrómica • Un axón puede conducir en ambas direcciones. • Cuando un PA se inicia en su parte media se establecen dos impulsos que transcurren en sentidos opuestos a causa de la conducción electrotónica. • En un animal vivo los impulsos solo se conducen en un sentido desde las uniones sinápticas o receptores a lo largo de los axones hasta el final, es decir ortodrómica. • La conducción sentido contrario se llama antidrómica.
- 160. AAxxoonnaall TTrraannssppoorrtt (fast & slow) (fast only)
- 162. Secuencia de fenómenos en la contracción del músculo esquelético • 1. descarga de la neurona motora • 2. liberación del neurotransmisor • 3. unión de la Ach con los receptores nicotínicos para acetilcolina • 4. aumento de la conductancia al sodio y al potasio en la membrana de la placa terminal • 5. generación del potencial de placa terminal • 6. generación del potencial de acción en las fibras musculares • 7. propagación de la despolarización hacia el interior a lo largo de los túbulos T • 8. liberación de Ca2+ de las cisternas terminales y difusión de este ión a los filamentos gruesos y delgados • 9. unión del Ca 2+ con la troponina C y se descubren los sitios de unión de la actina con la miosina • 10. formación de enlaces cruzados entre la actina y la miosina y deslizamiento de los filamentos delgados sobre los gruesos, lo que produce el acortamiento
- 163. Exitación
- 165. Mecanismo de deslizamiento de los filamentos en la contracción
- 166. Secuencia de fenómenos en la relajación del músculo esquelético • 1. el calcio se bombea al retículo sarcoplásmico • 2. liberación de calcio de la troponina • 3. cesa la interacción entre la actina y la miosina
- 167. Relaxation of Muscle Fiber
- 168. Secuencia de fenómenos en la contracción del músculo esquelético • 1. descarga de la neurona motora • 2. liberación del neurotransmisor • 3. unión de la Ach con los receptores nicotínicos para acetilcolina • 4. aumento de la conductancia al sodio y al potasio en la membrana de la placa terminal • 5. generación del potencial de placa terminal • 6. generación del potencial de acción en las fibras musculares • 7. propagación de la despolarización hacia el interior a lo largo de los túbulos T • 8. liberación de Ca2+ de las cisternas terminales y difusión de este ión a los filamentos gruesos y delgados • 9. unión del Ca 2+ con la troponina C y se descubren los sitios de unión de la actina con la miosina • 10. formación de enlaces cruzados entre la actina y la miosina y deslizamiento de los filamentos delgados sobre los gruesos, lo que produce el acortamiento
- 171. Clasificación de las neuronas • Las neuronas se clasifican desde el punto de vista morfológico en cuatro tipos principales según su forma y la distribución de sus proyecciones. • Las neuronas unipolares: Poseen una sola proyección y son raras en los vertebrados, salvo al principio del desarrollo embrionario. • Las neuronas bipolares: Poseen dos proyecciones que salen del soma, una sola dendrita y un solo axón. Las neuronas bipolares están localizadas en los ganglios vestibular y coclear y en el epitelio olfatorio de la cavidad nasal. • Neuronas seudounipolares: Poseen una sola proyección que sale del cuerpo celular, pero esta proyección se ramifica más tarde en una rama periférica y una central. La rama central entra en el SNC, y la rama periférica procede hacia su destino en el cuerpo. • Neuronas motoras. Poseen su cuerpo neuronal periférico
- 172. Las neuronas se clasifican también en tres grupos generales según su función: • Neuronas sensitivas (aferentes): Reciben estimulación sensitiva a nivel de sus terminaciones dendríticas y conducen impulsos hacia el SNC para su procesamiento. Las localizadas en la periferia del cuerpo vigilan los cambios en el ambiente, y las que se encuentran dentro del cuerpo vigilan el ambiente interior. • Neuronas motoras (eferentes): Se originan en el SNC y conducen sus impulsos hacia músculos, glándulas y otras neuronas. • Las interneuronas: Están localizadas dentro del SNC, funcionan como integradoras estableciendo redes de circuitos neuronales entre las neuronas sensitivas y motoras y otras interneuronas.
- 173. • Los nervios periféricos de los mamíferos están compuestos por muchos axones juntos en una envoltura fibrosa llamada perineuro
- 174. Tipo de fibra función Diámetro mM Vel. Cond. m/seg A a PROPIOCEPCIÓN, MOTORA, SOMÁTICA 12-20 70-120 MOTORA 5-12 30-70 b TACTO, PRESIÓN MUSCULARES 3-6 15-30 g MOTORA EN HUSOS d DOLOR , FRÍO, TACTO 2-5 12-30 B AUTÓNOMAS PREGANGLIONARES <3 3-15 C RAIZ DORSAL DOLOR, TEMP, MECANO RREC, RESPUESTAS REFLEJAS 0.4-1.2 0.5-2 POSGANGLIONARES 0.3-1.3 0.7-2.3 SIMPÁTICA SIMPÁTICAS
- 175. Clasificación numérica para neuronas sensoriales número origen Tipo de fibra Ia Huso muscular Aa Ib Órgano tendinoso de Golgi Aa II Huso muscular, terminación en ramillete, tacto, presión Ab III Receptores para dolor y frío, algunos receptores para tacto Ab IV Dolor temperatura y otros receptores C
- 176. Susceptibilidad de las fibras nerviosas A, B y C al bloqueo de la conducción. susceptibilidad +++ susceptible s ++ susceptible s +- susceptible s hipoxia B A C presión A B C Anestésicos locales C B A
- 177. • Las proteínas G son transductores de señales que llevan información desde el receptor hasta una o más proteínas efectoras. • Existe una gran superfamilia de receptores para numerosos fármacos destinatarios que interactúan con ciertas proteínas reguladoras heterotriméricas ligadas a GTP conocidas como proteínas G. • Los receptores acoplados a la proteína G (G protein-coupled receptors, GPCR) comprenden a los de varias aminas biógenas, eicosanoides y otras moléculas que envían señales a lípidos, péptidos hormonales, opiáceos, aminoácidos como GABA y muchos otros péptidos y ligandos proteínicos.
- 179. • Los efectores que son regulados por la proteína G comprenden enzimas como: • la adenililciclasa, • fosfolipasa C, • Fosfodiesterasas • También regulan canales de iones de la membrana plasmática selectivos para Ca2+ y K+. • Gracias a su número e importancia fisiológica, los GPCR constituyen objetivos muy utilizados para los fármacos; quizás la mitad de los fármacos que no son antibióticos están dirigidos hacia estos receptores, que constituyen la tercera familia más grande de genes en el ser humano.
- 180. • Forman una familia de proteínas caracterizadas por su interacción con guanosín trifosfato (GTP) conducente a la hidrólisis del nucleótido a guanosín difosfato (GDP). • Su nombre deriva la inicial de guanosina. • En la fisiología celular actúan como interruptores biológicos mediante la transducción de señales. • Un estímulo del exterior celular, un ligando por ejemplo, accede al receptor celular asociado a proteína G o GPCR desencadenado una cascada de actividades enzimáticas o segundos mensajeros como respuesta.
- 181. • Debido a su estructura molecular, las proteínas G se clasifican en dos tipos, heterotriméricas y monoméricas. • Las primeras, grandes o heterotriméricas, están constituidas por tres subunidades distintas, denominadas αβγ; se trata de proteínas ancladas a membrana, aunque no integrales de membrana. • Las segundas, pequeñas o monoméricas, con una única subunidad, se encuentran libres en el citosol y nucleoplasma.
- 182. Proteínas G
- 183. • Las proteínas G están integradas por subunidades cada una como un transmisor de señal metabólica especifica. • Se sabe que existe centenares de estas proteínas en los circuitos metabólicos del sistema nervioso, circulatorio y anabólico. • Este tipos de proteínas G tienen un papel importante en todas las células del organismo
- 184. • Estas proteínas acoplan a mas de 100 receptores distintos para diversas proteínas • como la adenilato ciclasa, la guanilato ciclasa, y algunos tipos de canales iónicos
- 185. • PROTEINAS G: – HETEROTRIMÉRICAS – Monomericas • CARACTERISTICAS COMUNES: – Transducen señales de membrana – Son interruptores moleculares: on/off y tiempo – Unen GTP, que es fundamental para su activación. – Hidrolizan GTP
- 186. • Las proteínas G grandes, o heterotriméricas, se sitúan en la membrana plasmática, a la cual están ligadas por sus subunidades α y γ mediante estructuras hidrofóbicas, de tipo ácido graso. • Queda una tercera subunidad, la β, que se asocia a la subunidad γ. • En conjunto, y en reposo, es decir, con GDP unido a la subunidad α, las tres subunidades se sitúan en algún lugar de la membrana biológica, poseyendo movimiento dada su fluidez.
- 187. • Cuando un GPCR recibe un estímulo y se activa a la proteína G, con la adquisición de GTP, que es hidrolizado posteriormente, ésta se disgrega en dos partes: • βγ, que puede ejercer funciones biológicas como la apertura de canales; y otra, • α, que activa cascadas de señalización celular, como por ejemplo mediante adenilato ciclasa y la generación de AMP cíclico.
- 188. Las proteínas G Unión a Estado activo nucleótidos de guanina Estado inactivo Trifosfato de guanosina GTP Di fosfato de guanosina GDP
- 189. Las proteínas G son heterotrimeros compuestas por las subunidades Alfa(con 39-46 kilodalton de peso molecular), Beta (37 kD) Gamma (8 kD).
- 190. • Las proteínas G se clasifican: • Si estimulan la actividad de la adenil ciclasa en alguna o varias de sus isoformas en Gs • si inhiben la actividad de dicha enzima en Gi y todas las subunidades alfa que activan distintas isoformas de fosfolipasa Cβ
- 191. Clases de proteínas G
- 192. LAS PROTEINAS G SE CLASIFICAN EN 3 GRUPOS 1.-Gs Activa a la AC 2.-Gi Que inhibe a la AC ,incluye a la transducina y a la gustducina 3.-Gq Que activa principalmente la cascada de fosfoinositidos , a las proteínas G12, se encuentran en diversos tipos de células incluido el sistema nervioso
- 194. El proceso de activación de proteínas efectoras inducidas por ligando puede explicarse según un modelo en tres fases 1 El receptor (azul claro) se encuentra en estado de reposo, así como el efector (rojo pálido). La proteína G se encuentra con todas sus subunidades asociadas y con GDP; por lo tanto, inactiva. 2 La unión de la hormona induce un cambio conformacional en el receptor, que se activa (azul oscuro). 3 El receptor activado se une a la subunidad Gβγ. 4 A consecuencia de dicha interacción, Gα cambia su conformación, se disocia el GDP y se adopta GTP, por lo que la proteína se encuentra activa. Gα se desplaza separándose del resto de subunidades aprovechando su independencia derivada de la posesión de anclas en la bicapa lipídica. 5 La hormona se disocia del receptor, por lo cual se inactiva (azul claro). Gα se une al efector y lo activa (rojo). El efector realiza una actividad catalítica. 6 La hidrólisis de GTP a GDP causa la disociación de Gα del efector y se reasocia con Gβγ. Todo el sistema queda en reposo, inactivo.
- 195. la activación de cascadas complejas de mensajeros intracelulares denominados secundarios ActivaLons a GGPsC, RG i, Gq Produciendo respuestas Regulación directa de canales iónicos Regulación de la fosforilacion de proteínas
- 196. • Las proteínas G monoméricas o pequeñas son GTPasas de masas moleculares entre 20 y 40 kDa Actúan como reguladoras de procesos claves, como : • la proliferación celular (p. ej. Ras), • tráfico de vesículas (p. ej. Rab) • la estructura del citoesqueleto (p. ej. Rho). • En el mismo polipéptido se presenta tanto la actividad GTPasa como la capacidad de reconocimiento de motivos estructurales en otras moléculas, siendo además muy móviles en el interior celular, sin poseer la restricción de encontrarse ligadas las membranas celulares. • Para regular su actividad GTPasa, existen también proteínas GEF y GAP, antes mencionadas. • Pertenecen a la superfamilia Ras en la que se han identificado más de 100 proteínas diferentes subdivididas en 5 subfamilias: Ras, Rho, Rab, Arf y Ran (Barandier et al., 2003), • En muchos casos, se encuentran solubles en el citosol
- 197. • Los genes codificantes de los distintos miembros de la familia Ras están muy conservados evolutivamente, lo que sugiere que estas proteínas son muy importantes en los procesos celulares. • Los genes ras se expresan en todos los tipos celulares y órganos, aunque existen diferencias en cuanto a expresión pre- y postnatal, y en ciertos tejidos adultos se expresan preferentemente uno u otro miembro de la familia. H-ras se expresa mayoritariamente en cerebro, músculo y piel; K-ras en estómago, pulmón y timo; mientras que N-ras se expresa en testículos y timo.
- 198. • Las proteínas Ras de mamíferos están localizadas en la cara interna de la membrana plasmática como homodímeros o heterodímeros (Santos et al., 1988). Para la unión de la proteína Ras a la membrana son necesarias una serie de señales moleculares que determinan la ruta por la cual esta proteína se ancla en la membrana y su localización en islotes lipídicos (rafts o caveolas, regiones ricas en colesterol) o en zonas desorganizadas de la membrana (Hancock et al., 1990). • la localización de Ras en la membrana también depende de estas modificaciones postraduccionales. Así, se sabe que H-Ras está localizada en islotes lipídicos, mientras que K-Ras se localiza en zonas desorganizadas de la membrana (Prior et al., 2001; Roy et al., 1999).
- 199. • Modificaciones postraduccionales y transporte de Ras a la membrana.(Adaptado de Rojas and Santos, 2002.)
- 200. RECEPTORES ACOPLADOS A PROTEÍNAS G
- 201. • Estos receptores celulares median respuestas a su interacción con diversas moléculas de señalización como lo son los neurotransmisores, neuropéptidos, hormonas, péptidos vasoactivos, aromatizantes, saborizantes, glucoproteínas y otros mediadores locales.
- 202. • de los receptores de esta clase tienen una estructura similar, esta consiste en una cadena polipeptídica simple con siete segmentos α-hélice trasmembranales que tienen una estructura tridimensional común (TM I-VII)
- 203. • estos dominios están unidos entre si por asas polipeptídicas tres intracelulares (i1-i3), • y tres asas extracelulares (e1-e3) una cuarta asa citoplasmática puede formarse cuando el segmento C- terminal se une a la membrana por atracción lipídica a la cadena de aminoácidos
- 204. • un segmento N-terminal glucosilado extracelular, el segmento C-terminal a nivel citoplasmático
- 205. • En cuanto a su estructura, los siete dominios transmembrana poseen dos características clave: • Orientación con el extremo amino terminal hacia el exterior y el carboxilo terminal hacia el interior. • Estructura de siete alfa-hélices transmembrana (H1 a H7), cuatro segmentos extracelulares (E1 a E4) y cuatro segmentos citosólicos (C1 a C4). El segmento carboxiterminal, el tercer bucle citosólico (C3) y, a veces, también el segundo (C4) están implicados en la interacción con la proteína G.
- 207. • De acuerdo a la homología de secuencia y similitud funcional, se clasifican en seis grandes grupos: • Clase A (o 1) (receptores semejantes a rodopsina) • Clase B (o 2) (receptores de de la familia secretina) • Clase C (o 3) (receptor metabotrópico de glutamato) • Clase D (o 4) (receptores fúngicos involucrados en la determinación sexual) • Clase E (o 5) (receptores de AMPc) • Clase F (o 6) (Frizzled/Smoothened, de las vías Wnt y hedgehog, respectivamente)
- 208. En cuanto a los efectores de las proteínas G, los de mamíferos pueden clasificarse de Clase GΑ Efector asociado Segundo mensajero Ejemplos de receptores Gsα Adenilil ciclasa Aumento de AMPc β-adrenérgico, glucagón, serotonina, vasopresina Giα Adenililciclasa, canal de K+ Disminución AMPc, cambio en el potencial de membrana α2-adrenérgico, muscarínico de acetilcolina Golfα Adenilil ciclasa Aumento AMPc Olfatorios Gqα Fosfolipasa C Aumento IP3, DAG α1-adrenérgico Goα Fosfolipasa C Aumento IP3, DAG Acetilcolina de células endoteliales GtΑ cGMP fosfodiesterasa Disminución cGMP Rodopsina de las células bastón acuerdo a los siguientes grupos
- 209. Principal Mecanismo de Acción Cuando no hay un ligando, el receptor está asociado a la proteína G. Cuando un ligando se une al receptor, lo cambia conformacionalmente: La Proteína G se separa del receptor y la subunidad α se une a un GTP. Al unir el GTP y sustituirlo por un GDP, las subunidades β y γ se separan de α . La unidad α + GTP produce efectos metabólicos diversos que no siempre se dan simultáneamente.
- 210. Principal Mecanismo de Acción – Activa la adenilato Ciclasa. – Activa la fosfolipasa C. – Provoca cambios en apertura y cierre de los canales iónicos.
- 211. Principal Mecanismo de Acción Seguidamente, se produce la separación de las subunidades desde la proteína G, en 2 componentes, la subunidad α y la βγ. Finalmente, al hidrolizar al GTP, la subunidad α se desactiva, restableciendo su unión con el dímero βγ y la proteína G queda lista para un futuro ciclo de activación.
- 212. Sistema Acoplado a la Adenilato Ciclasa • La AC es la enzima que sintetiza al 2º mensajero llamado AMPc que a partir de la hidrólisis del ATP, Y la enzima que degrada al AMPc recibe el nombre de fofodiesterasa (PDE).
- 213. Sistema Acoplado a la Adenilato Ciclasa • Cuando la AC se activa, su subunidad catalítica convierte al ATP en AMPc, y este segundo mensajero activa a la denominada proteína cinasa que propaga y amplifica las señales acopladas al sistema de la AC.
- 215. Sistema Acoplado a la Guanilato Ciclasa • Los sistemas celulares poseen un tipo de proteína cinasa llamada proteína cinasa G (PKG), dependiente de GMPc.
- 216. Sistema Acoplado a la Guanilato Ciclasa • La guanilato ciclasa es la que sisntetiza al 2º mensajero GMPc a partir de la hidrólisis del GTP, siendo la PKG homóloga a la PKA. • El aumento citoplásmico del GMPc activa a PKG uniéndose al domino regulador de la enzima y causando un cambio conformacional.
- 217. Sistema Acoplado a la Fosfolipasa C • Receptores asociados a proteínas Gq después de interactuar con su ligando, estimulan a la fosfolipasa C. • Lo que se traduce en un aumento en el recambio de fosfoinosítidos y la liberación de calcio de compartimentos intracelulares.
- 218. • La fosfolipasa C produce la catabolización de una molécula: el Fosfatidil Inositol, un fosfolípido de la membrana Fosfatidil inositol Inositol fosfato Diacilglicerol
- 219. Cuadro 7.1- Cuadro comparativo de las vías de transmisión a través de segundos mensajeros LOCALIZACIÓN CELULAR VÍA ADENILATO CICLASA (AC) Pasos generales VÍA DE LOS FOSFATO DE INOSITOL Espacio extracelular Adrenalina ¯ Inductor (Primer mensajero) ¯ Adrenalina ¯ Membrana plasmática Receptor b-adrenérgico ¯ Proteína Gs ¯ Adenilato ciclasa (AC) ¯ Receptor ¯ Transductor ¯ Amplificador ¯ Receptor a1-adrenérgico ¯ Proteína Gq ¯ Fosfolipasa C (PLC) ¯ Citosol ATP ¯ AMPc ¯ Proteinquinasa A (PKA) ¯ Fosforilación de Fosforilquinasas ¯ Glucógeno ® Glucosa Precursor Fosforilado ¯ Segundo mensajero ¯ Fosforilación de Proteinquinasas ¯ Fosforilaciónes enzimáticas ¯ Respuesta Celular PIP2 ¯ DAG - IP3 - Ca2+ ¯ Proteinquinasa C (PKC) ¯ Liberación de Ca2+ al citosol ¯ Vasoconstricción
- 221. • Uno que se une a proteínas G • Otro que se fosfolira durante el proceso de desensibilización del receptor
- 223. Las células poseen 2 tipos de GTP-asas • Monomérica y trimérica • Cada uno tiene la capacidad de unir trisfosfato de guanosina (GTP) y permite que se una ala subunidad alfa de la proteína G que a su ve z intercambia su GDP por un GTP
- 224. La union de GTP da lugar a que la subunidad alfa se disocie y esta se une con ciclasa de adenilato. Se da la activacion de la ciclasa de adenilato. Se desacopla el ligando del receptor ligado a la proteína G y el receptor regresa a su configuración original sin afectar la actividad de la subunidad alfa
- 226. La subunidad alfa hidroliza su GTP en GDP, se desprende de la ciclasa de adenilato y se une a otra vez a las subunidades Beta. Así la G1 Actúa en forma similar a la G5, pero se inhibe y no se produce cAMP Esto impide la fosforilación. Por eso la unión de un ligando particular a un receptor especifico puede activar o inactivar la célula según el tipo de proteína G que se acopla.
- 228. AMP CÍCLICO Y SU FUNCIÓN COMO SEGUNDO MENSAJERO El cAMP es una molécula de señalamiento intracelular que activa la cinasa de proteína dependiente de cAMP y se une en ella. La cinasa A se disocia en su componente regulador y 2 subunidades catalíticas activas.
- 230. • Fosforilan a otras enzimas en el citosol y por consiguiente inicia una cascada de fosforilaciones y su efecto es una respuesta especifica
- 231. • La transcripción de genes se dan por valores elevados de cAMP. • Se fosforila la cinasa A y por lo tanto se activa la proina de union de los elementos de respuesta de cAMP que estimula a los genes
- 232. • Cuando se encuentre cAMP en una concentración elevada suficiente da lugar a la reacción de la célula blanco
- 233. • El cAMP se degrada por la acción de las fosfodiesterasas de cAMP en 5- AMP