Fotosíntesis: proceso clave para la vida
•
0 gefällt mir•318 views

El documento describe el proceso de fotosíntesis en plantas. La fotosíntesis consta de dos fases: la fase luminosa, en la cual se captura la energía de la luz para producir ATP y NADPH a través de los fotosistemas I y II; y la fase de fijación de CO2, donde el ATP y NADPH se usan para fijar el CO2 y reducirlo a azúcares en el ciclo de Calvin. La energía de los fotones genera un gradiente protónico que permite la producción de ATP.

Empfohlen
Weitere ähnliche Inhalte
Was ist angesagt?
Andere mochten auch
Andere mochten auch (20)
Ähnlich wie Fotosíntesis: proceso clave para la vida
Ähnlich wie Fotosíntesis: proceso clave para la vida (20)
Fotosíntesis: proceso clave para la vida
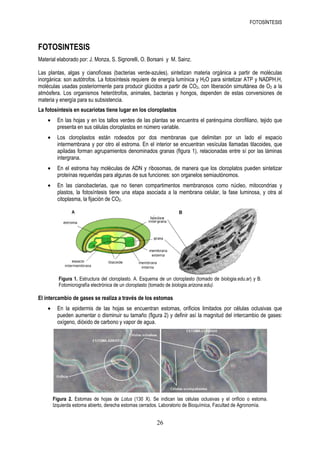
- 1. FOTOSÍNTESIS FOTOSINTESIS Material elaborado por: J. Monza, S. Signorelli, O. Borsani y M. Sainz. Las plantas, algas y cianofíceas (bacterias verde-azules), sintetizan materia orgánica a partir de moléculas inorgánica: son autótrofos. La fotosíntesis requiere de energía lumínica y H2O para sintetizar ATP y NADPH.H, moléculas usadas posteriormente para producir glúcidos a partir de CO2, con liberación simultánea de O2 a la atmósfera. Los organismos heterótrofos, animales, bacterias y hongos, dependen de estas conversiones de materia y energía para su subsistencia. La fotosíntesis en eucariotas tiene lugar en los cloroplastos • En las hojas y en los tallos verdes de las plantas se encuentra el parénquima clorofiliano, tejido que presenta en sus células cloroplastos en número variable. • Los cloroplastos están rodeados por dos membranas que delimitan por un lado el espacio intermembrana y por otro el estroma. En el interior se encuentran vesículas llamadas tilacoides, que apiladas forman agrupamientos denominados granas (figura 1), relacionadas entre sí por las láminas intergrana. • En el estroma hay moléculas de ADN y ribosomas, de manera que los cloroplatos pueden sintetizar proteínas requeridas para algunas de sus funciones: son organelos semiautónomos. • En las cianobacterias, que no tienen compartimentos membranosos como núcleo, mitocondrias y plastos, la fotosíntesis tiene una etapa asociada a la membrana celular, la fase luminosa, y otra al citoplasma, la fijación de CO2. Figura 1. Estructura del cloroplasto. A. Esquema de un cloroplasto (tomado de biologia.edu.ar) y B. Fotomicrografía electrónica de un cloroplasto (tomado de biologia.arizona.edu). El intercambio de gases se realiza a través de los estomas • En la epidermis de las hojas se encuentran estomas, orificios limitados por células oclusivas que pueden aumentar o disminuir su tamaño (figura 2) y definir así la magnitud del intercambio de gases: oxígeno, dióxido de carbono y vapor de agua. Figura 2. Estomas de hojas de Lotus (130 X). Se indican las células oclusivas y el orificio o estoma. Izquierda estoma abierto, derecha estomas cerrados. Laboratorio de Bioquímica, Facultad de Agronomía. 26
- 2. FOTOSÍNTESIS En la fotosíntesis se distinguen dos fases, la luminosa y la de fijación de CO2 Como cualquier proceso bioquímico la fotosíntesis se puede representar por una ecuación global, que en este caso resume una reacción de óxido-reducción en la que el H2O cede electrones (en forma de hidrógeno) para la reducción del CO2 a glúcidos (CH2O)n, con liberación de O2. CO2 + H2O luz O2 + (CH2O)n Fase luminosa En esta etapa, también llamada fotodependiente, porque se da sólo en presencia de luz, ocurren dos procesos bioquímicos necesarios para la síntesis de glucosa: la reducción de NADP a NADPH.H con los hidrógenos de la molécula de agua y la síntesis de ATP. • En la fase luminosa los pigmentos como las clorofilas a y b (figura 3), carotenos, xantofilas, ficoeritrinas y ficocianinas, que se encuentran asociados a la membrana tilacoidal, captan fotones y se excitan. Figura 3. Modelo de una molécula de clorofila. La región inferior de la molécula corresponde a una cadena hidrofóbica. En la parte superior se observa el anillo con dobles enlaces, y asociado a un Mg. Modificado de efn.uncor.edu • Los fotones, o cuantos de luz, son cantidades discretas de energía que se propagan como ondas. La luz visible corresponde a la región del espectro electromagnético comprendida entre 400 y 700 nm, región en la que absorben los pigmentos presentes en las plantas y cianobacterias. • Según su longitud de onda (λ), los fotones tienen diferente cantidad de energía, de manera que cuanto menor es λ, mayor es la energía, que decrece desde la región violeta a la roja (figura 4). Figura 4. Espectro electromagnético. Se representan las regiones del espectro y sus longitudes de onda. La longitud de onda está relacionada inversamente con la energía. Modificado de a-diba.net 27
- 3. FOTOSÍNTESIS La diversidad de pigmentos permite absorber fotones en todo el rango del espectro visible • Los pigmentos, con máximos de absorción en longitudes de onda características (figura 5), se agrupan en la membrana tilacoidal en estructuras llamadas antenas, formadas por unas 200 – 300 moléculas. Los distintos pigmentos absorben fotones con diferentes longitudes de onda, de manera que su diversidad le da a la planta la posibilidad de aprovechar todo el espectro visible (figura 5). Figura 5. Espectros de absorción de pigmentos fotosintéticos. El conjunto de pigmentos absorbe fotones en todo el rango visible del espectro. Estos pigmentos forman las antenas, mientras que el centro de reacción está formado sólo por clorofila. Tomado de monografías.com Los pigmentos de la antena se excitan con la energía de los fotones • Los pigmentos contienen electrones fácilmente excitables. Los electrones más externos pueden absorber la energía de un fotón y pasar a un estado excitado de mayor energía, y por lo tanto más inestable. Los electrones excitados se estabilizan al volver al estado basal, liberando la energía absorbida (figura 6). Figura 6. Esquema de la excitación de un electrón. Un electrón perteneciente a un orbital molecular puede ser excitado y pasar a otro orbital molecular de mayor energía, por el aporte externo de energía. En el ejemplo usado la energía proviene de un fotón (h.ν). El mismo electrón al volver al estado basal libera la energía absorbida (ε). 28
- 4. FOTOSÍNTESIS • De esta forma, la energía se transmite entre moléculas adyacentes en la antena, hasta alcanzar el centro de reacción (figura 7). Como la transmisión de la excitación electrónica acompaña la pendiente de energía, esta pasa de un pigmento cuyo pico de máxima absorción es menor que el del pigmento siguiente, es decir, con menor requerimiento de energía para excitarse. Figura 7. Representación de la antena y centro de reacción de un fotosistema. Un fotón excita un electrón de un pigmento que funciona como captador de energía, que se transfiere de una molécula de pigmento a otra, hasta llegar al centro de reacción. Cuando se excita la molécula de clorofila del centro de reacción se oxida: pierde un electrón. Modificado de recursos.cnice.mec.esm • La transferencia de energía desde la antena al centro de reacción se realiza con una pérdida mínima, en 10-10 seg. En este proceso de transferencia es determinante el ordenamiento preciso en la membrana de los tilacoides de los componentes de la antena y del centro de reacción. La clorofila del centro de reacción se oxida: pierde un electrón • Las antenas, centros de reacción y transportadores de electrones (figura 8) conforman los fotosistemas: el fotosistema I (PSI) o P700 (cuyo centro de reacción tiene el máximo de absorción a 700 nm) y el fotosistema II (PSII) o P680 (cuyo centro de reacción tiene un máximo de absorción a 680 nm). • Cuando un fotón excita a un pigmento antena del PSII, un electrón se eleva a un potencial más alto, y cuando vuelve al nivel basal la energía es transferida a otro pigmento antena y así sucesivamente hasta que se excita el centro de reacción P680, que pierde un electrón. El electrón que pierde el P680 es captado por la feofitina, una clorofila sin Mg. A partir de la feofitina una cadena de transporte electrónico lleva al electrón al centro de reacción del PSI (figura 8). El PSII sin el electrón queda como un radical catiónico (con carga +). • El PSII requiere de un electrón para llenar el hueco electrónico que se generó. En esto es clave el papel del agua, que mediante la fotólisis se descompone en 2H+ y 2 e- (figura 8). Estos últimos llegan, de a uno, al P680 oxidado (figura 8), mientras el oxígeno se libera a la atmósfera como O2. • Cuando un fotón excita a un pigmento antena del PSI, ocurre lo mismo que en la antena del PSII. Un electrón se eleva a un potencial más alto y comienza una transferencia de excitación entre pigmentos de la antena, de manera que la energía es transferida al centro de reacción P700. Este se oxida, y el electrón perdido es captado por un aceptor primario diferente al del PSII (figura 8). 29
- 5. FOTOSÍNTESIS • El electrón cedido por el P700 pasa por una secuencia de transportadores y después de una serie de reacciones redox, reduce al NADP a NADPH.H (figura 8). El NADPH.H aporta en la fase de fijación de CO2 el poder reductor necesario para reducir el CO2 a glucosa (figura 10). • Observar que ocurre un continuo flujo de electrones desde el agua hasta el NADPH.H, según se observa en el diagrama Z representado en la figura 8. En este flujo no cíclico de electrones ocurren reacciones redox, donde pierden gradualmente energía que es usada para generar un gradiente protónico y sintetizar ATP. • Cuando no es necesario el poder reductor ocurre el flujo cíclico de electrones. En este, los electrones del P700 llegan a la ferredoxina, y en lugar de derivar al NADP vuelven nuevamente al P680 (figura 8): el flujo cíclico permite producir ATP sin reducir al NADP. Figura 8. Diagrama Z: una representación del flujo de electrones a través de los fotosistemas. En el mismo diagrama se puede observar tanto el flujo cíclico de electrones como el no cíclico. Los electrones son excitados por la luz y pasan por una cadena transportadora de electrones a favor de un gradiente de energía hasta llegar al PSI, donde se había generado un hueco electrónico. Desde el centro de reacción del PSI sale un electrón por vez, que reduce al NADP, previo pasaje por una cadena de transportadores. Modificado de ecogenesis.com.ar La energía de los fotones genera un gradiente protónico que permite la producción de ATP • La energía liberada por los electrones entre el PSII y el PSI es utilizada para bombear protones contra gradiente electroquímico, a través de una membrana impermeable a H+, desde el estroma hacia el interior de los tilacoides (figura 9). • Este bombeo de protones desde el estroma produce alcalinización en este compartimento (pH 8) y acidificación del espacio intratilacoidal (pH 5). El potencial electroquímico que se genera a ambos lados de la membrana tilacoidal hace que los H+ pasen al estroma, a través de los canales protónicos CFo (figura 9). La energía liberada es usada para la conversión de ADP + Pi en ATP, proceso conocido como fotofosforilación. • La disposición de los transportadores de electrones y de la ATPasa en la membrana tilacoidal (figura 9), hace que el ATP y el NADPH.H se sinteticen del lado del estroma, donde serán consumidos en la fijación de CO2. • En mitocondrias y cloroplastos el mecanismo de síntesis de ATP se explica a través de la generación de un gradiente protónico. Una diferencia entre ambos es en la orientación del gradiente: en la mitocondria se establece desde el espacio intermembrana a la matriz (de afuera a adentro), con la 30
- 6. FOTOSÍNTESIS ATPasa dirigida hacia la matriz, mientras que en los cloroplastos es desde el espacio intratilacoidal hacia el estroma (de adentro hacia afuera), con la ATPasa dirigida hacia el estroma (figura 9). Figura 9. Esquema del proceso de fotofosforilación. La energía liberada por los electrones es usada para bombear protones desde el estroma al espacio tilacoidal. Esto crea un gradiente electroquímico cuya energía permite que los protones se muevan a favor del gradiente a través del complejo de CFo, y en la ATP sintetasa (CF1) se fosforila el ADP. Modificado de euita.upv.es Fase de fijación de CO2 La fijación de CO2 y su reducción a moléculas glucídicas (CH2O)n se da en el estroma del cloroplasto a través de reacciones reductivas, fuertemente endergónicas: el poder reductor (NADPH.H) y la energía (ATP) son producidos en la fase luminosa. Según el mecanismo de fijación de CO2 se establecen tres modelos fotosintéticos, el de las plantas C3, el de las C4 y el de las CAM (Crassulacean Acid Metabolism). Las más abundantes son las plantas C3, es decir, las que sólo presentan ciclo C3, porque tanto las plantas C4 como CAM presentan además un ciclo C3. Ciclo C3 o ciclo de Calvin: permite la conversión del CO2 en glúcidos • La energía y poder reductor generados en las reacciones fotoquímicas se usan para la conversión del CO2 en glúcidos en el Ciclo de Calvin (figura 10). Este ciclo ocurre en el estroma del cloroplasto, donde se encuentran las enzimas que catalizan la conversión de dióxido de carbono (CO2) en glúcidos (triosas y glucosa). • El CO2 ingresa a través de los estomas (figura 2) y se une al aceptor, la ribulosa 1,5 bifosfato, por acción de la enzima rubisco (ribulosa bifosfato carboxilasa oxigenasa). La rubisco constituye aproximadamente el 50% de las proteínas del cloroplasto y es la proteína más abundante en la naturaleza. • El primer producto estable después de la carboxilación es el ácido 3 fosfoglicérico (3PGA), un compuesto de 3 carbonos, al que se debe la denominación de ciclo C3. En “un ciclo” se fijan 6 moles de CO2 a 6 moles de ribulosa 1,5 bifosfato, y se forman 12 moles de 1,3PGA con un consumo de 12 moles de ATP (figura 10). • Para llegar a moléculas de glúcidos es necesaria la reducción del 1,3PGA a GA3P, lo que se logra con 12 moles de NADPH.H (figura 10): observar que la dirección de las reacciones son las mismas que las ocurridas en la glucogénesis. • El ciclo se cierra con la regeneración del aceptor, ribulosa 1,5P a partir de 10 moles de GA3P y el consumo de 10 moles de ATP: 10 GA3P x 3C = 30C (6 ribulosa 1,5P x 5C = 30C). 31
- 7. FOTOSÍNTESIS • Por otro lado los 2 moles de GA3P restantes pueden sintetizar glucosa en el cloroplasto y acumular almidón, o pueden ser transportado al citosol y ser usado en la glucólisis, o sintetizar glucosa y sacarosa, entre otros compuestos carbonados. • En períodos de oscuridad las células fotosintéticas se comportan como heterótrofas, y las células de tallos no fotosintéticos y raíces siempre se comportan como heterótrofas, de manera que dependen para su nutrición de moléculas glucídicas como la sacarosa, que llegan desde los tejidos fotosintéticos vía floema. Figura 10. Esquema del ciclo de Calvin. Se indica el número de moléculas que participan para sintetizar una molécula de glucosa: 6 CO2, 12 NADH.H y 12 ATP. Tomado de kambry.es La rubisco puede carboxilar u oxigenar a la ribulosa 1,5 P La enzima rubisco está formada por 16 subunidades, 8 mayores codificadas en el ADN del cloroplasto, y 8 menores codificadas en el ADN nuclear. • La rubisco tiene la propiedad de unir en el mismo centro catalítico al CO2 o al O2, es decir tiene dos sustratos, y cataliza la carboxilación o la oxigenación: de ahí su nombre ribulosa 1,5 bifosfato carboxilasa oxigenasa. • Cuando la enzima actúa como carboxilasa fija CO2 a la ribulosa 1,5P dando comienzo el ciclo C3: se asimila el carbono. Cuando la enzima actúa como oxigenasa, y oxigena a la ribulosa 1,5P, comienza un proceso conocido como fotorrespiración figura 11. • Si bien en la atmósfera hay un 21% de O2 y un 0.03% de CO2, la kM de la rubisco por el CO2 es 12 M, mientras que la kM por el O2 es 250 M. Esto hace que la asimilación del CO2 sea mayor a al incorporación de O2 a la ribulosa 1,5P, es decir, a la fotorrespiración. 32
- 8. FOTOSÍNTESIS Figura 11. Fotorrespiración y asimilación de carbono. La rubisco puede actuar como carboxilasa dando origen a la fijación de CO2, es decir a la asimilación de C, o como oxigenasa dando origen a la fotorrespiración. La función de la fotorrespiración es más evidente en plantas en condiciones de estrés La capacidad oxigenasa de la rubisco tiende a ser vista como un mecanismo inútil para la planta, porque desde el punto de vista de la eficiencia, no se aprecia una función útil para la planta: la fotorrespiración no conduce a la fijación sino a la pérdida de CO2, con gasto de ATP. • En condiciones normales de presión parcial de O2 y CO2 en atmósfera, hasta el 50% del carbono fijado por la fotosíntesis puede perderse por fotorrespiración. Así, este proceso reduce la eficiencia fotosintética en algunas plantas. • La importancia de este proceso es más comprensible en plantas en condiciones de estrés. A partir de resultados observados en plantas de tabaco, en las que se logró disminuir la velocidad de la fotorrespiración mediante ingeniería genética, se encontró que las plantas con niveles normales de fotorrespiración toleraron más la alta intensidad lumínica, respecto a las plantas con fotorrespiración disminuida. Ciclo C4: las plantas con esta estrategia minimizan la fotorrespiración • Plantas como Paspalum, maíz y caña de azúcar, entre otras, tienen una vía de captación de CO2 conocida como ciclo C4 que minimiza el proceso de fotorrespiración. Este ciclo ocurre en el citosol de las células del mesófilo, mientras que el ciclo C3, que ocurre seguidamente al ciclo C4, lo hace en los cloroplastos de las células de la vaina (figura 12). Figura 12. Corte trasversal de una hoja de Paspanum, una gramínea C4. Laboratorio de Botánica, Facultad de Agronomía. 33
- 9. FOTOSÍNTESIS • En las plantas C4 la PEP carboxilasa incorpora el CO2 a una molécula de fosfoenolpiruvado (PEP), de 3 carbonos, que se transforma en oxalacetato (OAA), de 4 carbonos (figura 13). El OAA se transforma en malato (o en aspartato), que pasa a la célula de la vaina donde hay cloroplastos con rubisco. En la célula de la vaina el malato es decarboxilado y rinde CO2 y una molécula de 3C (figura 13). • Mientras la molécula de 3C vuelve a la célula del mesófilo y regenera el PEP, con gasto de ATP, el CO2 es usado por la rubisco para carboxilar a la ribulosa 1,5P y dar comienzo al ciclo C3 (figura 13). • En las células de la vaina la concentración CO2 puede ser tres o cuatro veces mayor que su concentración en la atmósfera. De esta forma se evita buena parte de la actividad de oxigenasa de la rubisco, y la fotorrespiración no es evidente: las plantas C4 minimizan la fotorrespiración. • Esta estrategia hace a las plantas C4 más eficientes que las plantas C3, si se relaciona fotón absorbido con CO2 fijado, ambas en su condición óptima. En las plantas C4 la condición óptima se caracteriza por requerimientos de alta cantidad de luz, y elevada humedad y temperatura. Figura 13. Esquema general de la fijación de carbono por las plantas C4. En las células del mesófilo el CO2 se une al PEP por acción de la enzima PEPcarboxilasa (PEPc). El producto de esa reacción, de 4C, pasa a la célula de la vaina y abastece de CO2 al ciclo de Calvin (o ciclo C3) y permite la regeneración de PEP a partir del ácido piruvato(de 3C). Tomado de fisicanet.com.ar 34
- 10. FOTOSÍNTESIS Plantas CAM Como vimos hay tres tipos de plantas según el mecanismo fotosintético que presenten: las C3, las C4 y las CAM (Crassulacean Acid Metabolism). Ahora que están todos los mecanismos fotosintéticos descritos es importante hacer énfasis que, mientras que las plantas C3 sólo presentan el ciclo C3 para fijar CO2, las plantas C4 y las CAM presentan otro ciclo previo al C3, pero todas cuentan con un ciclo C3. • Las crasuláceas, cactáceas, bromeliáceas, y orquídeas son plantas con CAM. • La mayoría de las plantas son C3, de todas formas las plantas C4 y las CAM están presentes en la mayoría de los ecosistemas, si bien las plantas CAM (Figura 14) están adaptadas a condiciones extremas de temperatura y sequía, como ocurre en zonas desérticas. Figura 14. A la izquierda especies del género Crassula y Kalanchloe, a la derecha una cactácea. Prof. E. Marchesi, Laboratorio de Botánica Facultad de Agronomía. En las plantas CAM ocurren dos carboxilaciones en la misma célula, en diferente momento • Durante la noche, cuando los estomas están abiertos porque la temperatura desciende y la pérdida de agua por evapotranspiración es mínima, el CO2 ingresa a la planta y por acción de la enzima PEP carboxilasa se carboxila el PEP (Figura 15). Durante el día, en presencia de ATP y NADPH.H generados en la fase luminosa, ocurre la fijación de CO2 por la rubisco a través del ciclo C3: el ciclo C3 es común a todos los modelos fotosintéticos. • Las plantas CAM fijan el CO2 al PEP en la primera carboxilación y sintetizan oxalacetato, que se transforma en malato durante la noche. El malato que se almacena en las vacuolas de las células del parénquima es responsable del descenso del pH en las hojas durante la noche. El pH aumenta durante el día a medida que se consume el malato. • La descarboxilación del malato suministra CO2 a la rubisco, que cataliza la segunda carboxilación con la puesta en marcha del ciclo C3, que ocurre durante el día (Figura 15). El piruvato generado por decarboxilación del malato cierra el ciclo. • En las plantas CAM la fijación de CO2 está separada temporalmente en el día y la noche, mientras que en las plantas C4 la separación es espacial, es decir, en dos tipos de células, las del mesófilo y las de la vaina. • Las plantas CAM en general son facultativas, es decir tienen metabolismo CAM en determinadas condiciones ambientales y metabolismo C3 en otras. Si bien las plantas con esta modalidad de fijación de CO2 son características de regiones con climas extremos, su distribución geográfica es extensa, lo que probablemente logren por la flexibilidad metabólica que tienen para cambiar entre la modalidad C3 y CAM. Es más, es común que las plantas CAM funcionen como C3 en determinados momentos del día. 35
- 11. FOTOSÍNTESIS Figura 15. Esquema del proceso de fijación de CO2 por una planta CAM. Los estomas abiertos durante la noche permiten el ingreso de CO2, que se acumula como grupo carboxilo del oxalacetato. Durante el día la actividad de la fase luminosa genera ATP y NADPH.H, que permiten la reducción del CO2 aportado por el malato a través del ciclo C3. • Como la PEP carboxilasa y el CO2 liberado de la decarboxilación del malato están en el mismo compartimiento, si la enzima no estuviera regulada se generaría un ciclo fútil de carboxilación - decarboxilación. Por esto es necesario que la PEP carboxilasa esté regulada de tal manera que se evite que el CO2 liberado de la descarboxilación del malato sea nuevamente captado por el PEP para volver a para formar malato. Durante el día se evita esta situación porque la PEP carboxilasa está inhibida por malato, y durante la noche la PEP carboxilasa se activa, incluso en presencia de malato, por modificación covalente de la enzima: se fosforlila un residuo serina. Bibliografía Nelson y Cox, Lenhinger principios de bioquímica. Editorial Omega. Ediciones varias. Karp, C. Biología Celular y Molecular. Editorial McGraw-Hill Intermericana. 1996. http://www.alaquairum.net/fotosintesis.htm Links - Videos http://www.youtube.com/watch?v=BYWJHwF_yok&hl=es http://www.youtube.com/watch?v=HUo3xpcPTVc&feature=related http://www.youtube.com/watch?v=a8vFp_3vFEk&feature=related http://www.youtube.com/watch?v=mHU27qYJNU0 36