

Empfohlen
Weitere ähnliche Inhalte
Was ist angesagt?
Was ist angesagt? (16)
Andere mochten auch
Andere mochten auch (20)
Ähnlich wie Membrana struttura e funzione
Ähnlich wie Membrana struttura e funzione (18)
Membrana struttura e funzione
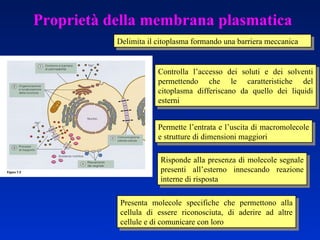
- 1. Proprietà della membrana plasmatica Delimita il citoplasma formando una barriera meccanica Controlla l’accesso dei soluti e dei solventi permettendo che le caratteristiche del citoplasma differiscano da quello dei liquidi esterni Risponde alla presenza di molecole segnale presenti all’esterno innescando reazione interne di risposta Permette l’entrata e l’uscita di macromolecole e strutture di dimensioni maggiori Presenta molecole specifiche che permettono alla cellula di essere riconosciuta, di aderire ad altre cellule e di comunicare con loro
- 3. Le teste polari dei fosfolipidi sono idrofile ( cercano l’acqua ) e sono quindi rivolte verso l’interno e l’esterno della cellula a contatto con le soluzioni acquose Le code apolari sono idrofobiche ( sfuggono l’acqua ) e sono rivolte verso l’interno della membrana E’ fondamentalmenete costituita da un doppio strato fosfolipidico spesso circa 5-8 nm, contenete una grande varità di proteine, una piccola percentuale di glucidi e, nelle cellule animali, numerose molecole di colestrolo
- 4. Modello a mosaico fluido
- 5. Il legami di tipo apolare che si formano tra le lunghe catene di acidi grassi e assieme alla loro tendenza a sfuggire il mezzo acquoso forniscono grande stabilità alla membrana fosfolipidica Il doppio strato di fosfolipidi
- 6. I TRE TIPI DI FOSFOLIPIDI: FOSFOGLICERIDI SFINGOLIPIDI STEROIDI FOSFOGLICERIDI (il C 3 del glicerolo è legato ad un gruppo fosfato, che, a sua volta, si lega ad un gruppo polare: Colina Etanolammina Glicerolo Inositolo: coinvolto nella trasduzione Serina Treonina Gli AG sono formati da 16, 18 0 20 atomi di carbonio. La maggior parte dei fosfogliceridi presenta un AG saturo lineare legato al C1 ed un AG insaturo con uno o più doppi legami , legato al C2
- 7. SFINGOLIPIDI Contengono , al posto del glicerolo, la sfingosina La sfingosina è un aminoalcool a lunga catena che contiene un gruppo amminico ed uno ossidrilico. il gruppo amminico è legato al gruppo carbossilico di un AG formando un ceramide Il gruppo ossidrilico può essere legato ad un ortofosfato, a sua volta esterificato con un aminoalcool: colina formando la sfingomielina GLICOSFINGOLIPIDI (CEREBROSIDI E GANGLIOSIDI)
- 9. Tra i fosfolipidi sono inserite molecole di colesterolo (in quantità variabile tra 0 e il 20% dei grassi). Il colesterolo dà particolari proprietà alle membrane fosfolipidiche impedendo ad esempio di irrigidirsi con l’abbassarsi della temperatura o di divenire troppo fluide al suo aumentare La percentuale di colesterolo può essere variata nel tempo in relazione alle esigenze della cellula
- 10. I singoli fosfolipidi si muovono di continuo navigando tra gli altri e permettendo alla membrana plasmatica di deformarsi Le teste idrofiliche del foglietto esterno sono diverse da quelle del foglietto interno. Per esempio, la fosfatidilserina, in una cellula integra, si trova rivolta soltanto verso il citosol, mentre viene esposta sulla superficie esterna quando la cellula muore.
- 11. PERCHE’ LE MEMBRANE CELLULARI DEVONO ESSERE FLUIDE? LA FLUIDITA’ PERMETTE ALLE PROTEINE DI MEMBRANA DI MUOVERSI LA FLUIDITA’ DELLA MEMBRANA PERMETTE LA FUSIONE DELLE MEMBRANE BIOLOGICHE
- 12. Fattori che influenzano la fuidità di membrana: TEMPERATURA : esiste una temperatura di fusione al di sopr della quale il doppio strato lipidico è allo stato fluido, mentre al di sotto i fosfolipidi di membrana vanno incontro ad un processo di gelificazione COMPOSIZIONE IN AG DEI FOSFOLIPIDI : La fluidità delle membrane cellulari diminuisce all’aumentare della lunghezza eal diminuire dell’insaturazione degli AG presentio nei fosfolipidi CONTENUTO DI COLESTEROLO : All’aumentare della temperatura la regione rigida e planare degli steroidi, ostacola il movimento reciproco dei fosfolipidi Al diminuire della temperatura la stessa regione impedisce le inetrazioni fta le code idrofobiche degli AG IL COLESTEROLO SI COMPORTA COME TAMPONE DI FLUIDITA’
- 13. LA SPECIFICITA’ FUNZIONALE ( adesione intercellualre, trasporto attivo, ricezioe di segnali di varia natura, riconoscimento.....) delle membrane dei diversi distretti cellulari è assicurata dalla componente proteica. Tutte le membrane, incluse quella plasmatica si originano a livello del Reticolo Endoplasmatico Rugoso È qui che le proteine di membrana vengono sintetizzate e associate alla membrana Per capire come questo avviene bisogna prima capire come la destinazione finale delle diverse proteine viene determinata
- 14. Le proteine vengono sintetizzate (utilizzando le informazioni degli RNA messaggeri) sia a livello dei ribosomi liberi (che si trovano nel citoplasma) sia a livello dei ribosomi associati alle membrane (reticolo endoplasmatico). La destinazione di queste proteine è differente a seconda di dove sono state prodotte L’ esatta destinazione delle diverse proteine dipende da una sequenza terminale che viene riconosciuta (e che poi viene staccata quando la proteina è stata trasportata alla sua destinazione)
- 16. Proteine di membrana - Intrinseche o integrali - Estrinseche o periferiche
- 17. Tra le proteine di membrana Intrinseche un caso particolare è rappresentato dalle proteine transmembranarie
- 18. PROTEINE DELLE MEMBRANE DEGLI ERITROCITI . Le due principali proteine di membrana dei globuli rossi, GLICOFORINA e PROTEINA della BANDA 3 , sono esempi di proteine transmembrana
- 20. La maggior parte delle proteine intrinseche può muoversi in orizzontale liberamente scorrendo dentro la membrana no no si si
- 21. Sulla superficie (specie quella esterna) della membrana si trovano anche dei glucidi per lo più legati ai lipidi ( glicolipidi ) o alle proteine ( glicoproteine ). Il complesso di catene glucidiche costituisce il GLICOCALICE. Per lo più essi servono da segnale di identità della cellula oppure reagiscono a specifiche sostanze dello spazio extracellulare
- 23. I principali tipi di proteine di membrana Canali ionici Recettore Enzima Ancoraggio del citoscheletro Esterno Esterno
- 24. CARATTERISTICHE DI ASIMMETRIA DELLA MEMBRANA MODIFICAZIONI DELLE MEMBRANE DELLE VARIE CELLULE : MIELINA MODELLO UNITARIO DELLE MEMBRANE
- 25. Caratteristiche di permeabilità della membrana plasmatica Se un solvente è più concentrato in un punto della soluzione esso tenderà a diffondersi lungo il gradiente di concentrazione , dalla parte più concentrata a quella meno concentrata La membrana plasmatica costituisce una barriera alla diffusione delle molecole verso l’interno e verso l’esterno della cellula Le cellule dei tessuti sono circondate dal liquido extracellulare che è molto diverso per composizione e concentrazione di soluti rispetto al citoplasma Diffusione
- 27. In sintesi la diffusione semplice attraverso una membrana fosfolipidica è un processo selettivo. Molecole piccole e apolari (quindi liposolubili e idrofobiche) diffondono molto velocemente (si dice anche la membrana è molto permeabile a queste sostanze). Al diminuire dell’affinità con i lipidi e all’ aumentare della dimensione , rallenta la velocità di diffusione. Gli ormoni sessuali per esempio (che sono steroidi e quindi grandi molecole affini al colesterolo) impiegano ore per penetrare in tutte le cellule Gli ioni infine diffondono così lentamente che per la maggior parte dei processi si può considerare che la membrana sia non permeabile a queste molecole
- 29. Diffusione facilitata Molecole che non passano attraverso lo strato fosfolipidico, possono tuttavia attraversare la membrana plasmatica lungo il loro gradiente di concentrazione utilizzando speciali proteine ( che sono specifiche per ciascuna molecola) Non è richiesta energia perché ciò che spinge le molecole è la differenza di concentrazione dai due lati della membrana Permettono selettivamente il passaggio di un tipo di ione (ad es il canale per il potassio) Sono utilizzati per far passare molecole più complesse (ad es il carrier per la cisteina) Canali ionici Carriers
- 31. I canali ionici sono di fondamentale importanza per il funzionamento della cellula nervosa Ogni attività del sistema nervoso, dalla percezione di uno stimolo sonoro, al richiamo di una memoria precedente, all’esecuzione di un movimento della mano dipendono dal gioco combinato dei canali ionici che si trovano sulla membrana plasmatica
- 32. Alcuni canali ( canali passivi ) non cambiano la loro permeabilità. Altri, i canali ad accesso variabile , possono variare la loro permeabilità in funzione di certi stimoli CANALE A CONTROLLO DI LIGANDOEXTRACELLULARE:Risponde ad un messaggero extracellulare CANALE A CONTROLLO DI LIGANDOINTRACELLULARE Risponde ad un messaggero intracellulare CANALE A CONTROLLO DI POTENZIALE :Risponde ad un cabiamento elettrico CANALE A CONTROLLO MECCANICO:Risponde ad uno stimolo meccanico
- 34. La pompa sodio potassio serve a mantenere le concentrazioni interne degli ioni. Essa è di fondamentale importanza per il funzionamento del neurone Per funzionare essa utilizza sia il meccanismo del trasporto accoppiato che l’energia dell’ ATP ENERGIA ( per il trasporto contro gradiente) ATP (es: pompa sodio-potassio) Co-trasporto (o trasporto accoppiato)
- 36. Ad ogni ciclo della pompa entrano due ioni potassio (K + ) ed escono tre ioni sodio (Na + ) Come effetto della pompa sodio-potassio, le concentrazioni di questi due ioni sono molto diverse all’interno e all’esterno della cellula Nel neurone ad esempio: - il potassio è 20 volte più concentrato all’interno della cellula rispetto all’esterno - il sodio è 10 volte più concentrato all’esterno
- 37. ENERGIA ( per il trasporto contro gradiente) ATP (es: pompa sodio-potassio) Co-trasporto (o trasporto accoppiato)
- 39. Endocitosi ed esocitosi Endocitosi: Una porzione del liquido extracellulare viene racchiusa a formare una vescicola Esocitosi: Il contenuto di una vescicola si riversa all’esterno mediante la fusione della membrana della vescicola con quella plasmatica Un meccanismo molto simile permette ai neuroni di liberare i loro neurotrasmettitori
- 40. Captazione o endocitosi mediata da recettori Endocitosi mediata da recettori: alcune specifiche proteine (es clatrina) rivestono esternamente porzioni di M. plasmatica specializzate, chiamate fossette rivestite . Quando tutti i recettori sono saturati dalla molecola che deve essere recuperata, la fossetta si invagina e si forma una vescicola Questo meccanismo è molto importante nelle sinapsi e serve al neurone pre-sinaptico per recuperare il neurotrasmettitore (ricaptazione)
- 41. RECETTORI DI MEMBRANA E COMUNICAZIONE CELLULARE
- 46. Le tre classi più grandi di recettori di superficie sono recettori collegati a canali ionici, a proteine G e a enzimi Sono coinvolti nella segnalazione Sinaptica rapida fra cellule eccitabili elettricamente Agiscono indirettamente nella regolazione dell’attività di una proteina bersaglio separata legata alla membrana (enzima o canale ionico) Quando sono attivati, o agiscono direttamente come enzimi o sono Direttamente associati ad enzimi che attivano
- 47. RECETTORI ASSOCIATI A PROTEINE G
- 48. Le proteine G trasmettono i loro segnali sulla superficie interna della membrana cellulare. Il processo comincia quando su un recettore si lega il corrispondente ormone o neurotrasmettitore, nell'esempio qui sopra l'adrenalina. Questo cambia la forma del recettore, che così si lega alla proteina G inattiva sul lato interno della membrana. Questo induce la proteina G ad espellere la sua molecola di GDP e a sostituirla con GTP. Il GTP fa cambiare la forma di un piccolo tratto di catena (mostrato qui in rosso), questo rende instabile la proteina G che si rompe in due frammenti.
- 49. La subunità alfa viene liberata col suo GTP legato, e si muove lungo la membrana finché trova l'enzima adenilil ciclasi. La piccola catena alfa si lega all'enzima e lo attiva. L'adenilil ciclasi attivato produce molto AMP ciclico che diffonde il segnale all'interno della cellula. Infine, il GTP nella subunità alfa attiva si rompe formando GDP, così la proteina G si può ricostruire per unione dei due frammenti che producono la forma trimera inattiva, pronta per eseguire un altro ciclo.
- 50. AMP CICLICO CALCIO La fosfolipasi attivata taglia PI(4,5)P2 generando due prodotti: inositolo 1,4,5-trifosfato (IP3) e diacilglicerolo.
- 51. IP3 è una piccola molecola solubile in acqua che lascia la membrana plasmatica e diffonde rapidamente nel citosol, dirigendosi verso il ER dove si lega ai canali del Ca++ aprendoli. Diacilglicerolo resta immerso nella membrana e va ad attivare una proteina chinasi C (PKC), Calcio-dipendente. L’aumento di Ca++ citosolico indotto da IP3 è essenziale per l’attivazione di PKC. PKC attivata fosforila proteine bersaglio tesuto-specifiche.
- 53. Le proteine G sono bersagli di alcune tossine importanti dal punto di vista medico. La tossina colerica , prodotta dal batterio che causa il colera. IPERSTIMOLAZIONE DELLE PROTEINE G O INIBIZIONE DELLE PROTEINE G La tossina della pertosse , prodotta dal batterio che causa la pertosse.
- 55. I SEGNALI EXTRACELLULARI SONO MOLTO AMPLIFICATI DALL’USO DI PICCOLI MEDIATORI INTRACELLULARI E DI CASCATE ENZIMATICHE.
- 56. SEGNALAZIONE TRAMITE RECETTORI DI SUPERFICIE COLLEGATI A ENZIMI Sono proteine transmembrana il cui dominio citosolico invece di associarsi ad una proteina G ha un’attività enzimatica intrinseca o si associa direttamente con un enzima. Le proteine segnale extracellulari che agiscono tramite recettori tirosina chinasi consistono di fattori di crescita (EGF,FGF,HGF,VEGF….) e ormoni. L’attivazione di un recettore tirosin chinasico dà luogo alla formazione di un complesso intracellulare per la trasmissione del segnale. Ogni tirosina fosforilata funge da sito specifico di legame per una proteina segnale diversa, e tutte propagano il segnale verso l’interno della cellula
- 58. Le varie vie di trasmissione del segnale interagiscono reciprocamente, mettendo in grado le cellule di produrre una risposta adatta a una combinazione di segnali complessa. Alcune combinazioni significano per la cellula che deve continuare a vivere, altri che deve riprodursi; in assenza totale di segnali quasi tutte le cellule si suicidano