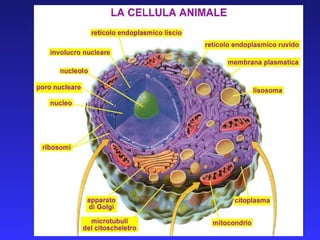
Smistamento delle proteine traffico vescicolare
•Als PPT, PDF herunterladen•
0 gefällt mir•5,170 views


Empfohlen
Weitere ähnliche Inhalte
Was ist angesagt?
Was ist angesagt? (20)
Andere mochten auch
Andere mochten auch (15)
Ähnlich wie Smistamento delle proteine traffico vescicolare
Ähnlich wie Smistamento delle proteine traffico vescicolare (20)
Smistamento delle proteine traffico vescicolare
- 2. RETICOLO ENDOPLASMATICO (RE) Reticolo endoplasmatico rugoso (RER) Reticolo endoplasmatico liscio (REL)
- 6. DETOSSIFICAZIONE ENZIMI CHE TRASFERISCONO L’OSSIGENO : OSSIGENASI (CITOCROMO P450)
- 7. COMPOSTI LIPOSOLUBILI COMPOSTI IDROSOLUBILI NETURALIZZAZIONE METILAZIONE DEMETILAZIONE IDROSSILAZIONE OSSIDAZIONE RIDUZIONE CONIUGAZIONE CON AA O ACIDI CONUIGAZIONE CON UN COMPOSTO MOLTO POLARE CHE NE PERMETTE IL PASSAGGIO NEL SANGUE E LA ELIMINAZIONE RENALE O L’IMMISSONE NELLA BILE E LA SUCCESSIVA ELIMINAZIONE CON LE FECI ALCUNI COMPOSTI DOPO I PROCESSI DI NEUTRALIZZAZIONE VENGONO TRASFORMATI IN C. MAGGIORMENTE TOSSICI : ALCUNI VELENI E PROCANCEROGENI
- 8. DETOSSIFICAZIONI O SINTESI PROTTETIVE REAZIONI CHIMICHE DI BIOTRASFORMAZIONE FASE I FASE II FASE I O FUNZIONALIZZAZIONE CONVERTONO LE SOSTANZE IN UN METABOLITA PIU ’ POLARE OSSIDAZIONE RIDUZIONE IDROLISI
- 9. FASE II CONIUGAZIONE CON UN SUBSTRATO ENDOGENO ACIDO GLUCURONICO ACIDO SOLFORICO ACIDO ACETICO AMINOACIDI
- 10. OSSIDAZIONI AVVENGONO NEL RE AD OPERA DI UN COMPLESSO E “drug metabolizing system” MONOOSSIGENASI CHE RICHIEDONO NADPH ED O 2 (CITOCROMI P450) RIDUZIONI AVVENGONO NEL CITOSOL E NEL REL DEL FEGATO E DI ALTRI TESSUTI IDROLISI AVVENGONO IN VARI TESSUTI AD OPERA DI ESTERASI E PROTEASI
- 11. ETANOLO Alcool deidrogenasi NAD dip. MEOS NADP dipendente Catalasi utilizza H 2 O 2 ACETALDEIDE ETANOLO ACETALDEIDE + NADH + H + ACETALDEIDE ACETATO +NADH + H + ACETATO ACETIL CoA INDUCIBILE DAL SUBSTRATO A.deidrogenasi
- 12. ACETALDEIDE LA SUA VELOCITA’ DI FORMAZIONE E’ SUPERIORE ALLA SUO CATABOLISMO ACCUMULO DANNI PRODOTTI DALL’ACETALDEIDE: INIBIZIONE SINTESI PROTEICA BLOCCO IMMISSIONE IN CIRCOLO LIPOPROTEINE DANNO MEMBRANE INIBIZIONE OX MITOCONDIALE DANNO CITOSCHELETRO
- 13. FASE II CONIUGAZIONE CON UN SUBSTRATO ENDOGENO ACIDO GLUCURONICO ACIDO SOLFORICO ACIDO ACETICO AMINOACIDI
- 14. GLUCURONAZIONE ACIDO GLUCURONICO+FENOLI O ACIDI AROMATICI: COMPOSTI MENO TOSSICI E PIU ’ SOLUBILI MOLTI FARMACI SUBISCONO CONIUGAZIONE GLUC. I GLUCURONIDI POSSONO ESSERE SCISSI E RIACQUISTARE IL LORO POTERE TOSSICO (B-GLUCURONIDASI EPATICHE O DAI BATTERI PRESENTI NELLA VESCICA: IN QUESTO CASO LA PRESENZA DEI COMPOSTI TOSSICI NELLA VESCICA PUO ’ DARE LUOGO A TUMORI : DERIVATI DELL’ANILINA)
- 15. ACETILAZIONE ELIMINAZIONE DI SOSTANZE TOSSICHE CONTENENTI GRUPPI AMINICI : SULFAMIDICI CONIUGAZIONE SOLFORICA AVVIENE NEL FEGATO E NELLA MUCOSA INTESTINALE FENOLO E ACIDO SOLFORICO LE SOSTANZE SONO PIU ’ SOLUBILI DEI FENOLI E QUINDI PIU ’ FACILMENTE ELIMINABILI COME I GLUCURONIDI POSSO ESSERE RIPRISTINATI O NEL FEGATO O NELLA VESCICA
- 16. TRASFORMAZIONI METABOLICHE CON AUMENTO DELLA TOSSICITA’ METABOLIZZAZIONE DEI VELENI CON FORMAZIONE DI COMPOSTI PIU ’ TOSSICI O PER DEGRADAZIONE DEL COMPOSTO O PER COMBINAZIONE CON SOSTANZE PRODOTTE NELL’ORGANISMO (SINTESI LETALE) CCl 4, FLUOROACETATO ETIONINA
- 18. LE PROTEINE SINTETIZZATE DEVONO POI PASSARE LA MEMBRANA DELL’ORGANULO BERSAGLIO: TIPI DI TRASPORTO TRASLOCATORI PROTEICI (mitocondri, reticolo endoplasmatico, perossisomi) PORI NUCLEARI (nucleo) TRASPORTO VESCICOLARE (Golgi, membrana, lisosomi, spazio extracellulare)
- 19. SINTESI DI PROTEINE INTEGRALI DI MEMBRANA PROTEINE DEI SISTEMI DI ENDOMEMBRANE PROTEINE DI SECREZIONE IN UNA CELLULA EUCARIOTE VENGONO SINTETIZZATE OLTRE 10.000 PROTEINE DIVERSE, CIASCUNA DELLE QUALI DEVE RAGGIUNGERE LA PROPRIA CORRETTA DESTINAZIONE. ANOMALIE NELLO SMISTAMENTO DELLE PROTEINE SONO ALLA BASE DI IMPORTANTI PATOLOGIE UMANE
- 24. VIA SECRETORIA: LO SMISTAMENTO INIZIA DURANTE LA TRADUZIONE
- 25. VIA CITOPLASMATICA : LO SMISTAMENTO INIZIA DOPO IL COMPLETAMENTO DELLA TRADUZIONE
- 28. LA VIA SECRETORIA: IPOTESI DEL SEGNALE: PRESENZA DI UN CODICE DI INDIRIZZO A LIVELLO DELLE PROTEINE. BLOBEL 1999: PREMIO NOBEL PER LA MEDICINA SEGNALE N-TERMINALE COSTITUITO 16-30 AMINOACIDI CON UN GRUPPO DI 6-12 AMINOACIDI IDROFOBICI, SPESSO PRECEDUTO DA UNO O PIÙ AMINOACIDI A CARICA POSITIVA. TALE SEQUENZA PUO’ ESSERE ANCHE INTERNA E NON APPENA ESCE DAL RIBOSOMA NE INNESCA L’ATTACCO AL RE ED IL MOVIMENTO DEL PEPTIDE NASCENTE ALL’INTERNO DELLE CISTERNE, ATTRAVERSO UN CANALE ACQUOSO PROTEICO DELLA MEMBRANA
- 30. La sequenza N-terminale è riconosciuta: da un complesso proteico NAC (Nascent Associated Complex) che le protegge da altre possibili interazioni da un complesso ribonucleico particella SRP (particella di riconoscimento del segnale) cosituita da una molecola di RNA (7S) e da sei proteine diverse. SRP lega un recettore per SRP presente sulla membrana del RE
- 33. Le tappe sopra descritte sono regolate dal legame e dall’idrolisi del GTP: Sia SRP che il suo recettore sono proteine G
- 35. NON APPENA IL POLIPEPTIDE NASCENTE ENTRA NELLE CISTERNE DEL RE, SI TROVA AD INTERAGIRE CON UNA MOLTITUDINE DI ENZIMI LOCALIZZATI SULLA PARTE E NEL LUME LA PEPTIDASI DEL SEGNALE (PROTEINA INTEGRALE LOCALIZZATA IN PROSSIMITA’ DEL TRASLOCONE ) RIMUOVE LA PORZIONE N-TERMINALE CONTENENTE LA SEQUENZA SEGNALE LA OLIGOSACCARILTRANSFERSI (PROTEINA INTEGRALE LOCALIZZATA IN PROSSIMITA’ DEL TRASLOCONE ) , AGGIUNGE CARBOIDRATI NEL LUME SONO PRESENTI ANCHE PROTEINE CHAPRON CHE CONSENTONO IL CORRETTO RIPIEGAMENTO SONO PRESENTI ANCHE DISOLFUROISOMERASI CHE CATALIZZANO LA FORMAZIONE DI PONTI DISOLFURO FRA I RESIDUI DI CISTEINA. DI FONDAMENTALE IMPORTANZA PER LA STABILITA’ DELLE PROTEINE PRESENTI SULLA SUPERFICIE CELLULARE EXTRACELLULARE O PER LE PROTEINE DI SECREZIONE
- 36. RETICOLO ENDOPLASMATICO : LA PORTA DI INGRESSO PER IL PERCORSO BIOSINTETICO DELLA CELLULA Il reticolo endoplasmatico è il compartimento membranoso più esteso della cellula. Sulla sua membrana possono aderire molti ribosomi (13 milioni) negli epatociti. Rappresenta anche la sede di smistamento delle proteine Destinate: al Golgi, ai lisosomi ed allo spazio extracellulare: viene considerato la porta di ingresso per il percorso biosintetico della cellula.
- 37. L’ambiente del lume favorisce il corretto ripiegamento e l’assemblaggio delle proteine e la divisione fra le proteine di secrezione, le proteine lisosomiali e le altre proteine neosintetizzate. La segregazione delle proteine neosintetizzate nel lume del RER, le sottrae dal citosol e consente che siano modificate e spedite verso la loro destinazione finale, sia fuori alla cellula che all’interno degli organuli.
- 38. SINTESI DELLE PROTEINE INTEGRALI DI MEMBRANA Le proteine integrali di membrana, tranne quelle dei mitocondri e dei cloroplasti, vengono sintetizzate sul RER. Queste proteine vengono trasferite al RER co-traduzionalmente, con lo stesso meccanismo descritto per la sintesi delle proteine di secrezione, ma presentano due regioni: la sequenza di inizio del trasferimento e la sequenza di arresto del trasferimento
- 39. la sequenza di inizio del trasferimento e la sequenza di arresto del trasferimento sono costituite da aa idrofobici che, transitando nel traslocatore inducono un suo cambiamento conformazionale: quando il traslocone (a forma di clessidra) incontra la sequenza Stop-transfer , si apre e rilascia la proteina
- 42. TUTTE LE PROTEINE TRANSMEMBRANA, MONO E MULTIPASSO PRESENTI NEL GOLGI, NEI LISOSOMI, NELLA MAMBRANA PLASMATICA, MA ANCHE QUELLE DEL LUME DEL GOLGI E DEI LISOSOMI, SEGUONO QUESTA VIA CHE INIZIA NEL RE E PROSEGUE AI VARI COMPARTIMENTI ATTRAVERSO UN PROCESSO DI TRASFERIMENTO MEDIATO DA VESCICOLE. QUANDO UNA MEMBRANA SI MUOVE DA UN COMPARTIMENTO ALL’ALTRO LA SUA COMPOSIZIONE VIENE MODIFICATA AD OPERA DEGI ENZIMI IVI RESIDENTI LA MAGGIOR PARTE DELLE PROTEINE DI MEMBRANA DEL RE VIENE MODIFICATA DALL’ATTACCO COVALENTE DI UNA O PIU’ CATENE OLIGOSACCARDICHE COSITUENDO COSI’ LE GLICOPROTEINE BIOSINTESI DELLE MEMBRANE NEL RE
- 43. ASIMMETRIA DELLE MEMBRANE L’ASIMMETRIA VIENE STABILITA INIZIALMENTE NEL RE QUANDO LIPIDI E PROTEINE VENGONO INCORPORATI IN MODO DIVERSO NEI DUE STRATI TALE ASIMMETRIA VIENE MANTENUTA QUANDO LA MEMBRANA PASSA DA UN COMPARTIMENTO ALL’ALTRO PER GEMMAZIONE E PER FUSIONE I COMPONENTI SITUATI SULLA SUPERFICIE CITOSOLICA DEL RE, SARANNO LOCALIZZATI SULLA SUP CITOSOLICA DELLE VESCICOLE DI TRASPORTO, SULLA SUP CITOSOLICA DEL GOLGI, E SULLA SUPERFICIE INTERNA (CITOPLASMATICA) DELLA MEMBRANA PLASMATICA. LO STESSO PER I COMPONENTI DELLA PARTE LUMINALE DELLA MEMBRANA
- 44. SINTESI DEI LIPIDI DI MEMBRANA LA MAGGIOR PARTE DEI LIPIDI E’ SINTETIZZATA NEL RE. ECCEZIONI: SLINGOMIELINE, GLICOLIPIDI: RE E POI GOLGI ALCUNI LIPIDI DI MITOCONDRI E CLOROPLASTI: NELLA SEDE STESSA GLI ENZIMI SONO LOCALIZZATI NELLA MEMBRANA DEL RE I LIPIDI INTEGRATI NELLA PORZIONE CITOSOLICA DEL RE VENGONO TRASPORTATI AL GOLGI ED ALLA MEMBRANA PLASMATICA. NEL PERCORSO VENGONO MODIFICATI
- 45. LA MAGGIOR PARTE DELLE PROTEINE DI MEMBRANA DEL RE VIENE MODIFICATA DALL’ATTACCO COVALENTE DI UNA O PIU’ CATENE OLIGOSACCARDICHE COSITUENDO COSI’ LE GLICOPROTEINE IMPORTANZA FUNZIONALE IMPORTANZA NEL FOLDING GLICAZIONE DELLE PROTEINE
- 47. Funzione della glicosilazione delle proteine La glicosilazione avviene per più motivi. Innanzitutto perché una proteina glicosilata raggiunge un f olding corretto e, in questo modo, può esplicare la propria funzione. Inoltre la glicosilazione protegge dall'attacco di proteasi ed aumenta la solubilità della molecola proteica che viene dunque stabilizzata in tutti gli aspetti. Infine il meccanismo glicosidico permette lo svolgimento del controllo di qualità Il principio di riconoscimento avviene sulla base della presenza o meno di un particolare residuo di glucosio sulla struttura glicosidica. La maggior parte delle proteine che vengono glicosilate, nelle cellule eucariotiche, sono destinate a diventare proteine di membrana: le catene di zuccheri vanno a formare infatti il glicocalice che circonda il plasmalemma (membrana plasmatica).
- 48. La N-glicosilazione vede l'aggiunta di una catena glucidica standard a livello dell'atomo di azoto di una catena laterale di asparagina ad opera di glucosiltransferasi. La N-glicosilazione ha inizio nel reticolo endoplasmatico rugoso a carico di una catena peptidica ancora in corso di traduzione.
- 49. La prima fase consiste nel trasferimento di una catena di 14 zuccheri (2 di N-acetilglucosammina, 3 di glucosio e 9 di mannosio) ad un residuo laterale di asparagina.
- 50. L'oligosaccaride è assemblato all'interno del reticolo endoplasmatico a partire da singoli carboidrati, ed è trasferito da uno speciale enzima (la glicosiltransferasi) da una molecola di dolicolfosfato (molecola incorporata nella membrana del RE) alla proteina, come singolo elemento
- 51. I primi 7 zuccheri: 5 mannosi e 2 residui di N Acetil Glucosamina sono traferiti nel citosol. Il dolico subisce flippaggio e ruota all’interno del RE Dove vengono aggiunti i restanti zuccheri: 4 mannosi e 3 glucosi. Completato lo zucchero viene trasferito al residuo di asparagina ed Il dolicolo puo’ essere esportato sul versante citosolico
- 52. Mutazioni che determinano la totale assenza della N-glicosilazione, causano la morte dell’embrione prima dell’impianto. Mutazioni che determinano la parziale alterazione della N-glicosilazione, causano gravi disordini ereditari. Negli organismi più evoluti l’oligosaccaride di base subisce una serie di modifiche nel RE come la rimozione di 2 glucosi. Questo è un passaggio fondamentale nella vita della glicoproteina neosintetizzata, che viene valutata da un sistema di controllo di qualità, prima di passare nel compartimento successivo della via biosintetica.
- 54. Il processo si ripete fino a quando la proteina non raggiunge il folding corretto. Se la proteina non è in grado di raggiungere un folding corretto, viene distrutta
- 55. Le proteine misfolded vengono esportate attraverso un meccanismo di «traslocazione inversa» nel citoplasma dove vengono degradate dai proteasomi
- 58. Ubiquitina
- 59. La proteina poli-ubiquitinata si lega al cappuccio del proteasoma La catena di ubiquitina viene rimossa ed il polipeptide denaturato ad opera della subunità beta del proteasoma mediante l’energia fornita da ATP
- 60. In certi casi le proteine misfolded si originano con velocità superiore rispetto a quella di esportazione nel citoplasma. Si attiva la risposta UPR (Unfolded protein response) Sensori proteici localizzati nel RE monitorizzano la concentrazione delle misfolded. In condizioni normali i sensori sono inattivi perché legati a chaperon (BiP) un aumento della concentrazione di unfolded determina il reclutamento di BiP e la attivazione dei sensori che dimerizzano e si auto fosforilano innescando una serie di segnali come:
- 61. La regolazione di geni che codificano per: chaperoni proteine coinvolte nel trasporto delle misfolded fuori dal RE ubiqutine La fosforilazione di eIF2, fattore di inizio della sintesi proteica, che si inattiva; determinando un rallentamento nella sintesi proteica
- 62. Controllo di qualità della sintesi di proteine
- 63. Le proteine che hanno subito sia la glicosilazione che la prima modificazione vengono trasportate, tramite vescicole, all'apparato del Golgi. Qui subiscono una sequenza ordinata di importanti cambiamenti e smistate verso le loro destinazioni definitive. In questo modo la cellula può regolare il trasporto di proteine, lipidi e carboidrati di nuova sintesi anche all’esterno (esocitosi). Smistamento delle proteine dal RE al Golgi, lisosomi, membrana e spazio extracellulare
- 65. L’apparato di Golgi E’ costituito da un insieme di cisterne appiattite, impilate le une sulle altre e a tratti fenestrate. Ogni cisterna è formata di membrane simili a quelle del RE, disposte a delimitare uno spazio di circa 10nm, che si allarga nelle regioni laterali (30-50nm).
- 66. Ogni pila può contenere da 3 a 20 cisterne. L’apparato di Golgi è un organulo strutturalmente e funzionalmente polarizzato La faccia cis è in genere rivolta verso il nucleo ed adiacente al RE; la faccia trans è rivolta verso la membrana plasmatica della cellula. La parte intermedia costituisce le cisterne mediali Il lato cis rappresenta la faccia immatura forma la rete cis del Golgi:CGN Il lato trans corrisponde alla faccia matura forma la rete trans del Golgi:TGN
- 67. La polarità dell’apparato di Golgi Nell’apparato di Golgi si distinguono tre compartimenti diversi ciascuno dotato di specifici enzimi ed adibito ad una determinata fase della elaborazione delle glicoproteine giunte dal RER. I tre compartimenti sono detti cis , mediano e trans .
- 69. La differenza fondamentale rispetto alla glicazione nel RE è la specificità. Nel reticolo endoplasmatico la glicosilazione è un evento "seriale", che non varia al variare del substrato. Nel Golgi ogni specifica proteina viene riconosciuta e modificata in base alla futura funzione. Si possono verificare rimozioni o aggiunte di singoli zuccheri o di catene più lunghe. La specificità delle singole catene glucidiche è il meccanismo utilizzato dalla cellula per lo smistamento delle proteine alle varie sedi di destinazione (lisosomi, membrana, perossisomi ). Completamento della glicosilazione delle glicoproteine iniziata nel RER
- 71. FUNZIONI DEL COMPLESSO DI GOLGI Nel Golgi si completa la sintesi della componente oligosaccaridica delle glicoproteine, iniziata nel RER
- 72. O-glicosilazione La O-glicosilazione è un processo altamente specifico, che non vede l'aggiunta "seriale" di carboidrati alla proteina in processazione. Si svolge completamente nell'apparato del Golgi, dove zuccheri vengono legati al peptide a livello dell'atomo di ossigeno delle catene laterali di serina o treonina. L'aggiunta riguarda un singolo carboidrato alla volta; solitamente il numero di zuccheri legati durante questo processo è limitato a pochi residui.
- 73. Dal reticolo trans Golgi gemmano vescicole con destino diverso
- 75. Sono state distinte diverse categorie di vescicole rivestite in base: alle proteine che costituiscono il loro rivestimento, al loro ruolo nel traffico cellulare ed al loro aspetto in microscopia
- 76. I tre principali tipi di vescicole sono : vescicole rivestite da COPII (spostano i materiali in avanti dal RE al Golgi) vescicole rivestite da COPI (spostano il materiale in senso retrogrado dal Golgi al RE) vescicole rivestite da CLATRINA (spostano il materiale dal TGN verso i lisosomi, gli endosomi ed i vacuoli. Spostano i materiali anche dalla membrana plasmatica ai compartimenti citoplasmatici lungo la via endocitotica e sono coinvolte nel traffico dagli endosomi ai lisosomi)
- 77. Le vescicole lisosomali che si staccano dal RER sono vescicole rivestite di clatrina
- 78. Clatrina : complesso proteico a 3 braccia (triskelion) formato da tre polipeptidi più grandi (heavy) e da tre più piccoli (light) in grado di polimerizzare e formare strutture esagonali a gabbia
- 79. LA FORMAZIONE ELLE VESCICOLE DERIVA DALLA POLIMERIZZAZIONE DELLA CLATRINA. CHE NECESSITA DELLA PRESENZA DI ALTRE MOLECOLE DI MEMBRANA: ADAPTINE IN GRADO DI RICONOSCERE IL CARGO DA TRASFERIRE LA POLIMERIZZAZIONE FA SOLLEVARE LA MEMBRANA ED ACCRESCERE LA VESCICOLA CHE SI STACCA DALLA MEMBRANA MEDIANTE L’INTERVENTO DI UNA PROTEINA LA DINAMINA. LE VESCICOLE MIGRANO NEL CITOSOL UTILIZZANDO ANCHE IL CITOSCHELETRO E SI FONDONO CON LE MEMBRANE DEGLI ORGANULI BERS AGLIO
- 80. IL RICONOSCIMENTO DELL’ORGANULO BERSAGLIO E’ MEDIATO DALLE PROTEINE SNARE (SNAP Receptor) E DALLE GTPasi Rab LE SNARE v-SNARE t-SNARE SONO PROTEINE FIBROSE ANCORATE ALLE MEMBRANE CAPACI DI RCONOSCIMENTO RECIPROCO. TALE RICONOSCIMENTO DETERMINA L’AVVICINAMENTO DELLE DUE MEMBRANE E LA FUSIONE SUCCESSIVA LE GTPasi Rab SONO IMPLICATE SIA NEL TRASFERIMENTO DELLE VESCICOLE LUNGO I MICROTUBULI , CHE NEL RICONOSCIMENTO DELLE SNARE
- 81. Il rivestimento di clatrina si disassembla, lasciando la vescicola nuda, ma dotata della sua vSNARE La vSNAre riconosce la tSNARE specifica e determina l’attacco della vescicola sulla membrana bersaglio Le SNARE, con il contributo di altre proteine, mediano la fusione delle due membrane e poi si liberano per un nuovo ciclo operativo
- 82. Le SNARE, oltre a fornire specificità nell’indirizzamento, contribuiscono anche alla fusione della membrana delle vescicole con quella del bersaglio. La specificità delle SNARE in una determinata vescicola sembra dipendere dal contenuto di questa o meglio dalle caratteristiche del recettore di carico posto sulla sua membrana.
- 83. Fusione delle membrane Dopo la fusione, le SNARE avvolte vengono separate ad opera di una proteina detta NSF che utilizza ATP. Proteine accessorie intervengono per favorire la fusione dei foglietti lipidici.
- 84. Dal reticolo trans del Golgi originano i lisosomi
- 85. I lisosomi Le proteine lisosomiali giungono a questo compartimento entrando inizialmente nel RE (grazie ad una sequenza di indirizzamento) a livello del Golgi vengono etichettate per poter essere indirizzate ai lisosomi. Questo indirizzamento è si basa sulla loro glicazione ad opera di un mannosio (fosforilato in posizione 6)
- 86. Il segnale mannoso-fosfato delle idrolasi lisosomali Produzione e smistamento degli enzimi lisosomali Nel compartimento cis il Golgi separa i futuri enzimi lisosomali dalle proteine destinate ad essere secrete
- 87. L’enzima GlcNAc fosfotransferasi riconosce specificamente le idrolasi lisosomali
- 89. Endocitosi Pinocitosi Fagocitosi Endocitosi mediata da recettori
- 90. L’endocitosi mediata da recettori costituisce una via d’accesso specifica nella cellula animale LDL: low density lipoprotein. Rappresenta la forma di trasporto del colesterolo alle cellule Difetti che alterano l’espressione o il funzionamento dei recettori di LDL sono alla base dell’aterosclerosi.
- 91. Endocitosi mediata da recettori nel ciclo della transferrina
- 92. GLI ENDOSOMI Attraverso il processo endocitotico prendono origine gli endosomi che internalizzano i materiali dall’esterno all’interno del citoplasma. Il comparto endosomico funge da centrale di smistamento della via endocitotica diretta verso l’interno. L’ambiente acido dell’endosoma induce molti recettori a liberarsi del loro bagaglio molecolare e ritornare nel loro dominio di membrana. Le molecole internalizzate negli endosomi sono prevalentemente destinate ai lisosomi per essere digerite.
- 93. I materiali destinati alla degradazione arrivano al lisosoma seguendo vie diverse
- 95. Dal trans Golgi originano vescicole di esocitosi
- 96. I segnali possono essere di sequenza di zone unite nella struttura
- 98. IL RITORNO AL RER ALCUNE PROTEINE DAL GOLGI VENGONO RIPORTATE AL RER CON UN TRASPORTO RETROGRADO
- 100. VIA CITOPLASMATICA : LO SMISTAMENTO INIZIA DOPO IL COMPLETAMENTO DELLA TRADUZIONE
- 101. ASSUNZIONE POST-TRADUZIONALE DI PROTEINE DA PARTE DI MITOCONDRI NUCLEO PEROSSISOMI CLOROPLASTI
- 103. Il trasferimento delle proteine richiede la presenza di complessi traslocatori TOM TIM dotati di una porzione di riconoscimento della porzione di indirizzamemto della proteina e di una porzione che costituisce il vero canale idrofilo per il passaggio della proteina La sequenza segnale ha la conformazione di un’alfa elica con una superficie basica
- 106. Hsp70 guida l’importazione delle proteine
- 107. Trasporto mitocondriale: proteine di membrana
- 108. Trasporto nucleare
- 109. TRASPORTO NUCLEARE Una delle caratteristiche più peculiari della membrana nucleare è costituita dalla presenza di aperture specializzate, chiamate PORI NUCLEARI
- 110. Ogni poro è costituito da un canale cilindrico che si estende attraverso le due membrane, permettendo la comunicazione tra citosol e nucleoplasma. Le due membrane sono fuse tra di loro a livello di ogni singolo poro formando un canale tappezzato da una complicata struttura proteica , il COMPLESSO DEL PORO Il complesso del poro nucleare ha un diametro di 80-100 nm.
- 111. Il complesso del poro (NPC) contiene più di 100 diversi tipi di subunità proteiche. Le subunità assumono una organizzazione ottagonale, protrudono da entrambi i lati dell’involucro nucleare. I NPC contengono al loro interno una struttura detta trasportatore, responsabile probabilmente del movimento delle macromolecole attraverso la membrana nucleare. Altre proteine collegano le otto subunità , dette anche bracci. Ci sono poi proteine che ancorano il NPC alle membrane ed altre che formano una specie di gabbia nel lato del nucleosoma
- 115. L’ATTRAVERSAMENTO DEL COMPLESSO DEL PORO DA PARETE DI MOLECOLE CON MASSA SUPERIORE A 50kDa E’ MEDIATO DALLA SEQUENZA DI LOCALIZZAZIONE NUCLEARE (NLS): brevi sequenze di aminoacidi carichi positivamente come lisina ed arginina
- 117. Ran GTPase dirigono il trasporto ALCUNE MOLECOLE ENTRATE NEL NUCLEO NON VI RIMANGONO (PROTEINE CHE REGOLANO LA TRASCRIZIONE) MA PER ESSERE INATTIVATE DEVONO ESSERE ESPORTATE SEQUENZE DI ESPORTZIONE NUCLEARE NES SONO RICONOSCIUTE DA RECETTORI DI ESPORTAZIONE
- 118. Il legame con Ran-GTP induce il rilascio del cargo
Hinweis der Redaktion
- Sistema piu esteso di membrane cellulari; forma un complesso sistema si sacchi appiattiti (cisterne) , vescicole e tubuli. I componenti del sistema sono tra loro intercomunicanti. Varia a seconda del tipo cellulare: ed es negli oociti è assente e si sviluppa gradualmente nell’embrione. RER ribosomi REL no Anche il rapporto tra RER e REL varia a seconda del tipo cellulare.
- Le dimensioni del REL possono variare considerevolmente in base alla funzione della cellula (ES: cellule del Leydig produce steroidi e possiede molto REL) Il rel è coinvolto in numerosi processi elenco. degli steroidi biosintesi dei lipidi di membrana (trasportati ai vari siti dalle vescicole o da proteine scambiatrici di fosfolipidi)
- IL REL è dotato di pompe specifiche per il ca++ che permettono di accumularlo in grandi conc nel suo lume. In seguito a specifici stimoli il ca++ puo essere rilasciato nel citoplasma e mediare diversi fondamentali meccanismi come la trasduzione del segnale stimolo nervoso contrazione muscolare.
- È il sistema di membrane più esteso della cellula eucariotica e, serve da punto di ingresso per proteine destinate ad altri organelli. Le proteine destinate al Golgi, agli endosomi, ai lisosomi come anche alla superficie cellulare, passano tutte dal citosol al RE Due tipi di proteine traslocano dal citosol al RE: Proteine idrosolubili Proteine destinate a una collocazione transmembrana
- Fig. 5.49
- Fig. 5.49
- Figg. 5.50 e 5.51
- Fig. 5.35
- Fig. 5.38
- Fig. 5.30